

A Requirement for Argonaute 4 in Mammalian Antiviral Defense
- PMID: 32049003
- PMCID: PMC7039342
- DOI: 10.1016/j.celrep.2020.01.021
A Requirement for Argonaute 4 in Mammalian Antiviral Defense
Abstract
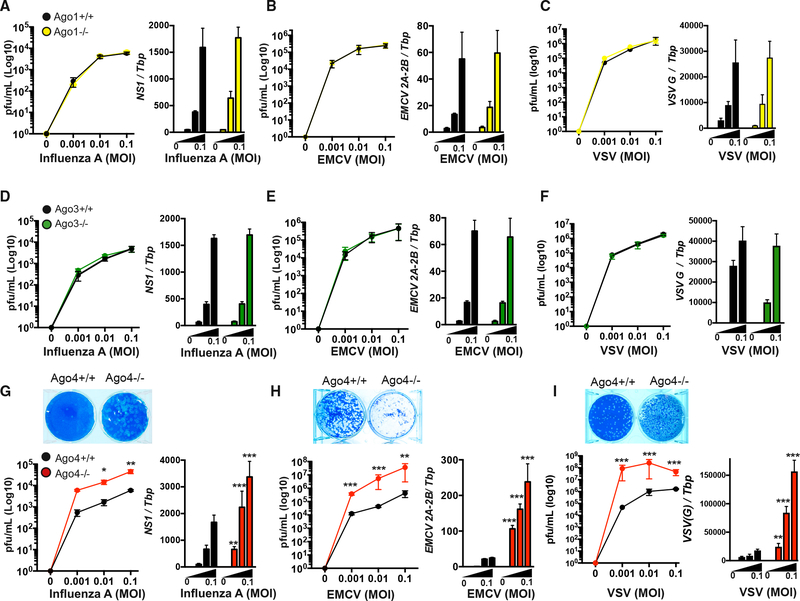
While interferon (IFN) responses are critical for mammalian antiviral defense, induction of antiviral RNA interference (RNAi) is evident. To date, individual functions of the mammalian RNAi and micro RNA (miRNA) effector proteins Argonautes 1-4 (AGO1-AGO4) during virus infection remain undetermined. AGO2 was recently implicated in mammalian antiviral defense, so we examined antiviral activity of AGO1, AGO3, or AGO4 in IFN-competent immune cells. Only AGO4-deficient cells are hyper-susceptible to virus infection. AGO4 antiviral function is both IFN dependent and IFN independent, since AGO4 promotes IFN but also maintains antiviral capacity following prevention of IFN signaling or production. We identified AGO-loaded virus-derived short interfering RNAs (vsiRNAs), a molecular marker of antiviral RNAi, in macrophages infected with influenza or influenza lacking the IFN and RNAi suppressor NS1, which are uniquely diminished without AGO4. Importantly, AGO4-deficient influenza-infected mice have significantly higher burden and viral titers in vivo. Together, our data assign an essential role for AGO4 in mammalian antiviral defense.
Keywords: Argonaute; MAVS; RNAi; antiviral immunity; influenza; interferon; macrophages; microRNA; vsiRNA.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures




References
-
- Ayllon J, and García-Sastre A (2015). The NS1 protein: a multitasking virulence factor. Curr. Top. Microbiol. Immunol 386, 73–107. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, and Hannon GJ (2001). Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409, 363–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
