

GLP-1 receptor agonists synergize with DYRK1A inhibitors to potentiate functional human β cell regeneration
- PMID: 32051230
- PMCID: PMC9945936
- DOI: 10.1126/scitranslmed.aaw9996
GLP-1 receptor agonists synergize with DYRK1A inhibitors to potentiate functional human β cell regeneration
Abstract
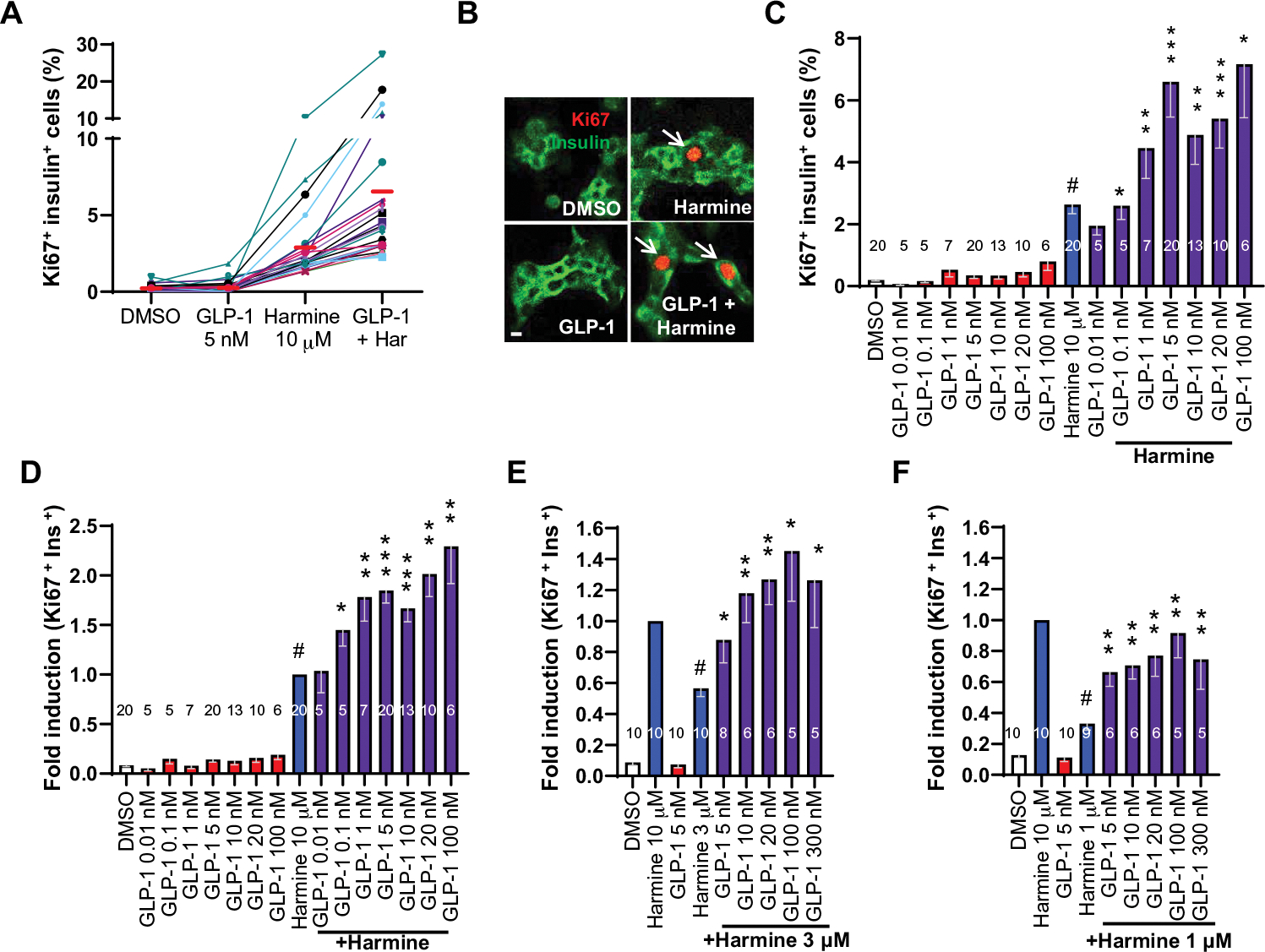
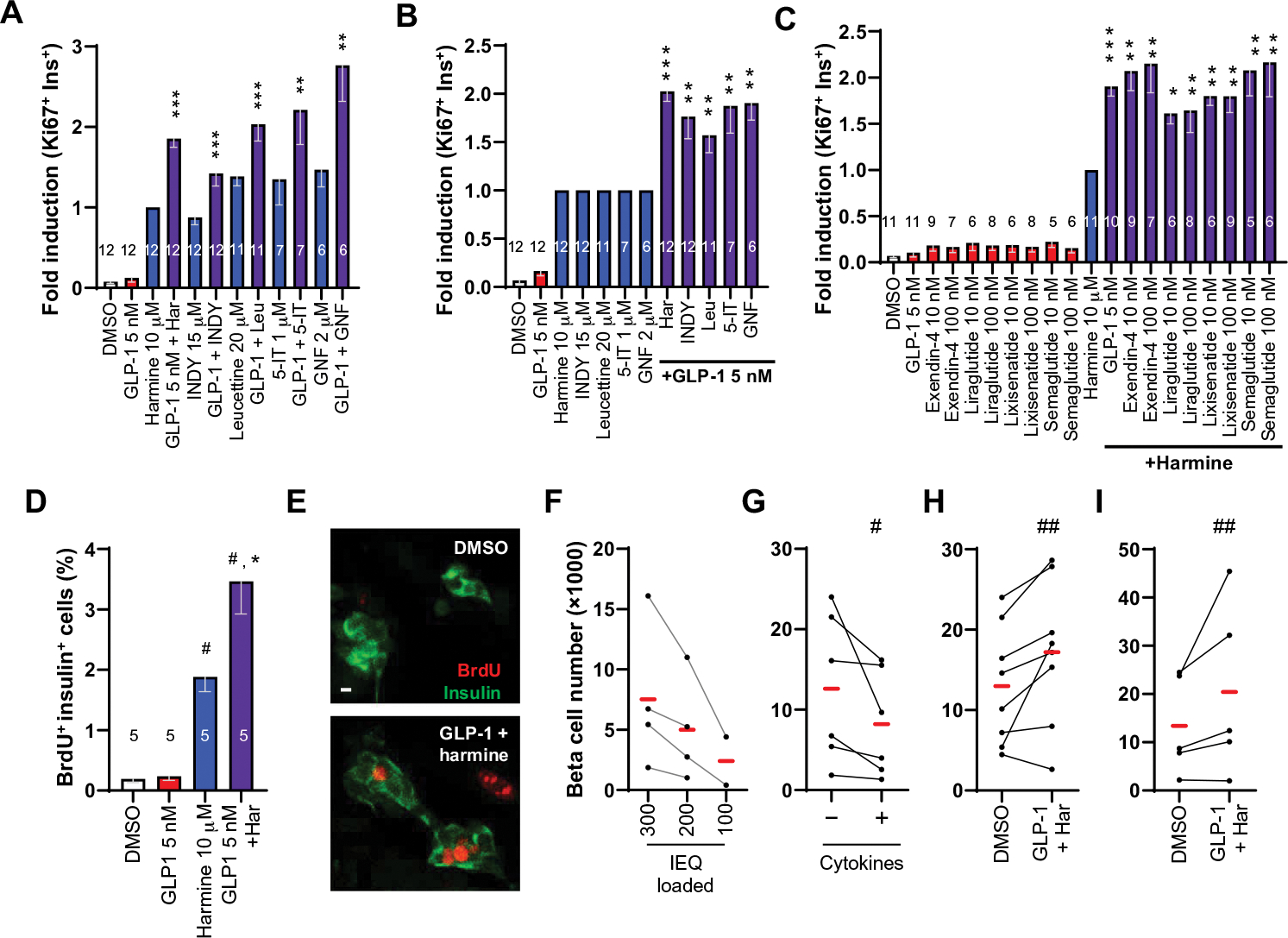
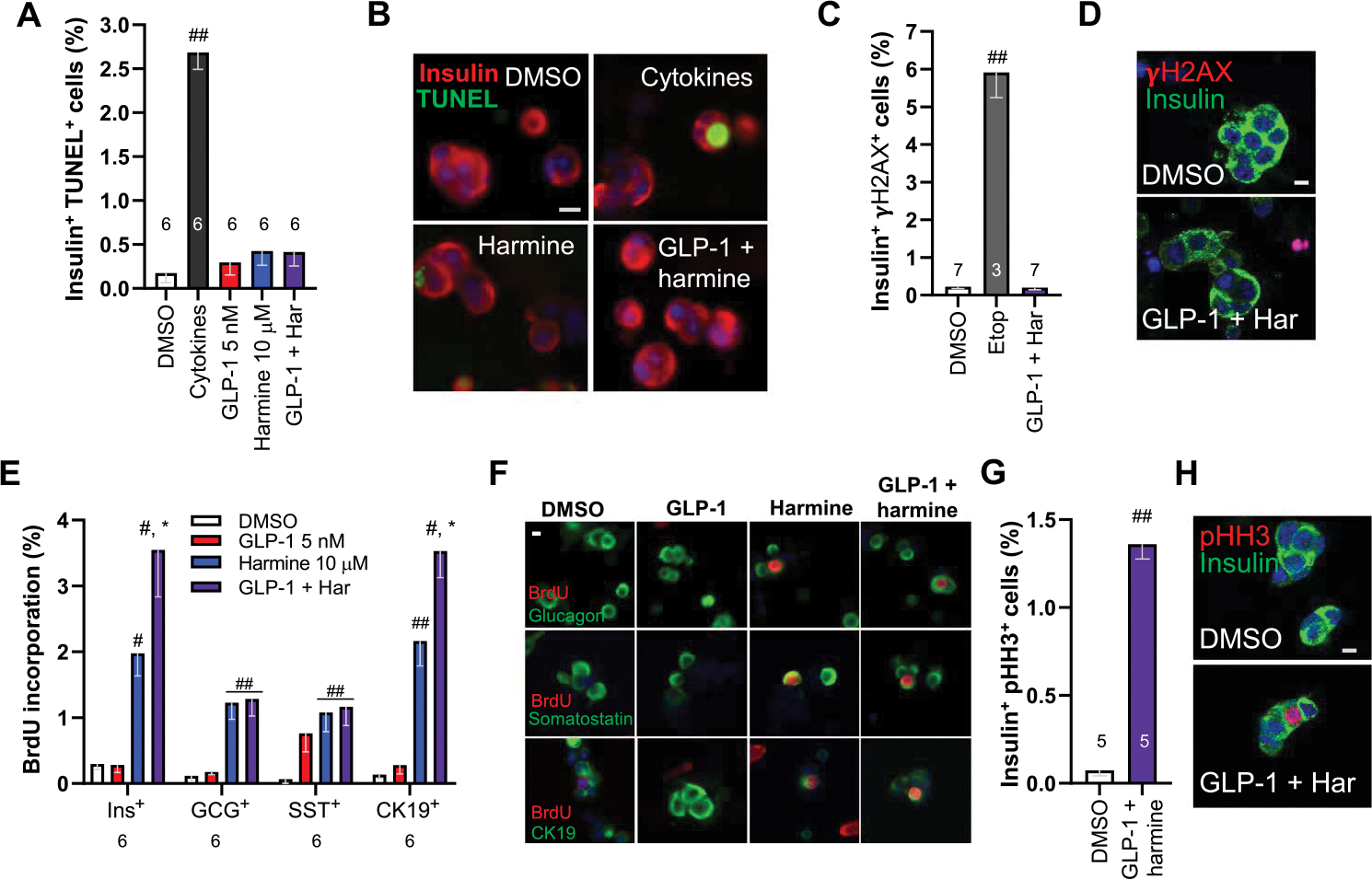
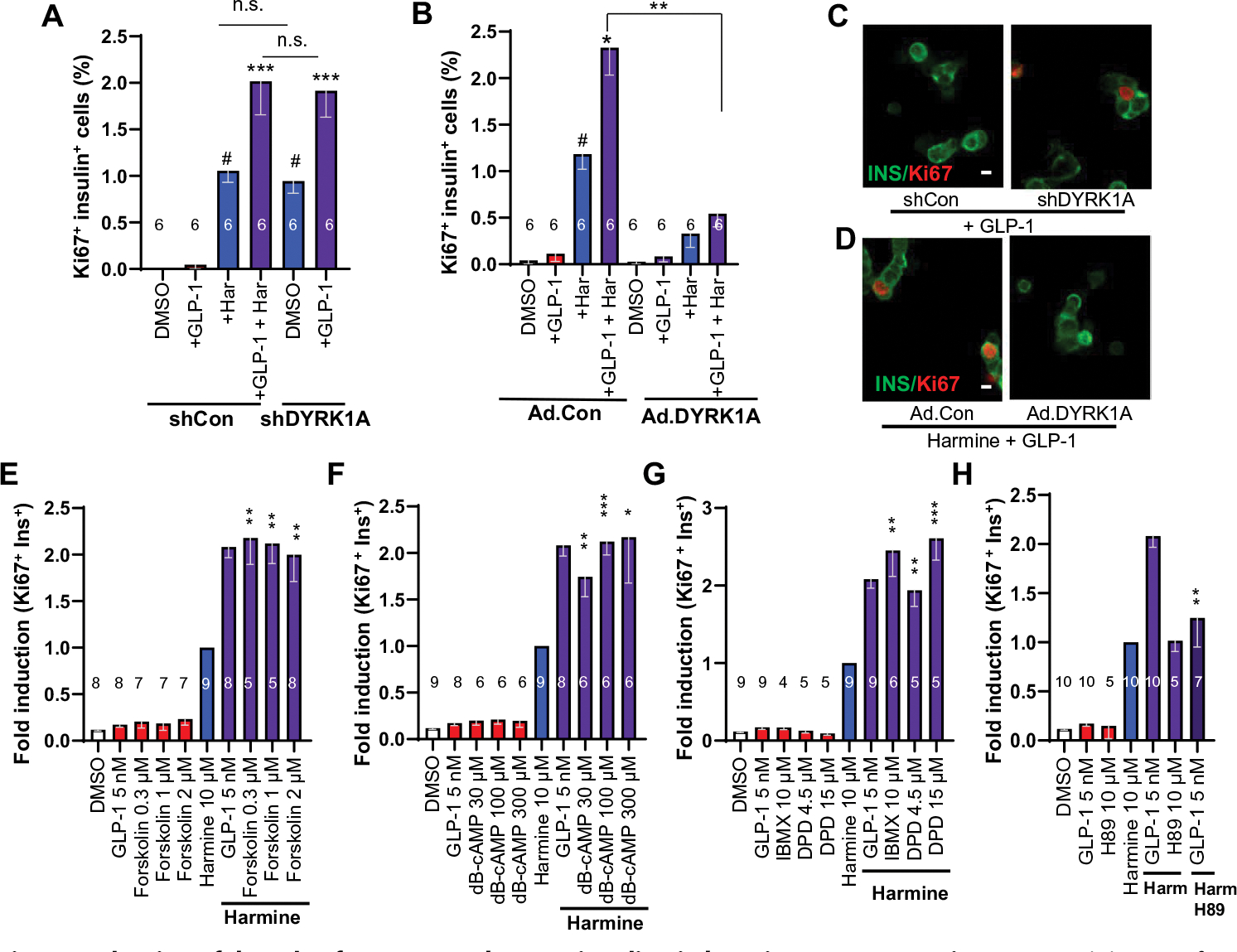
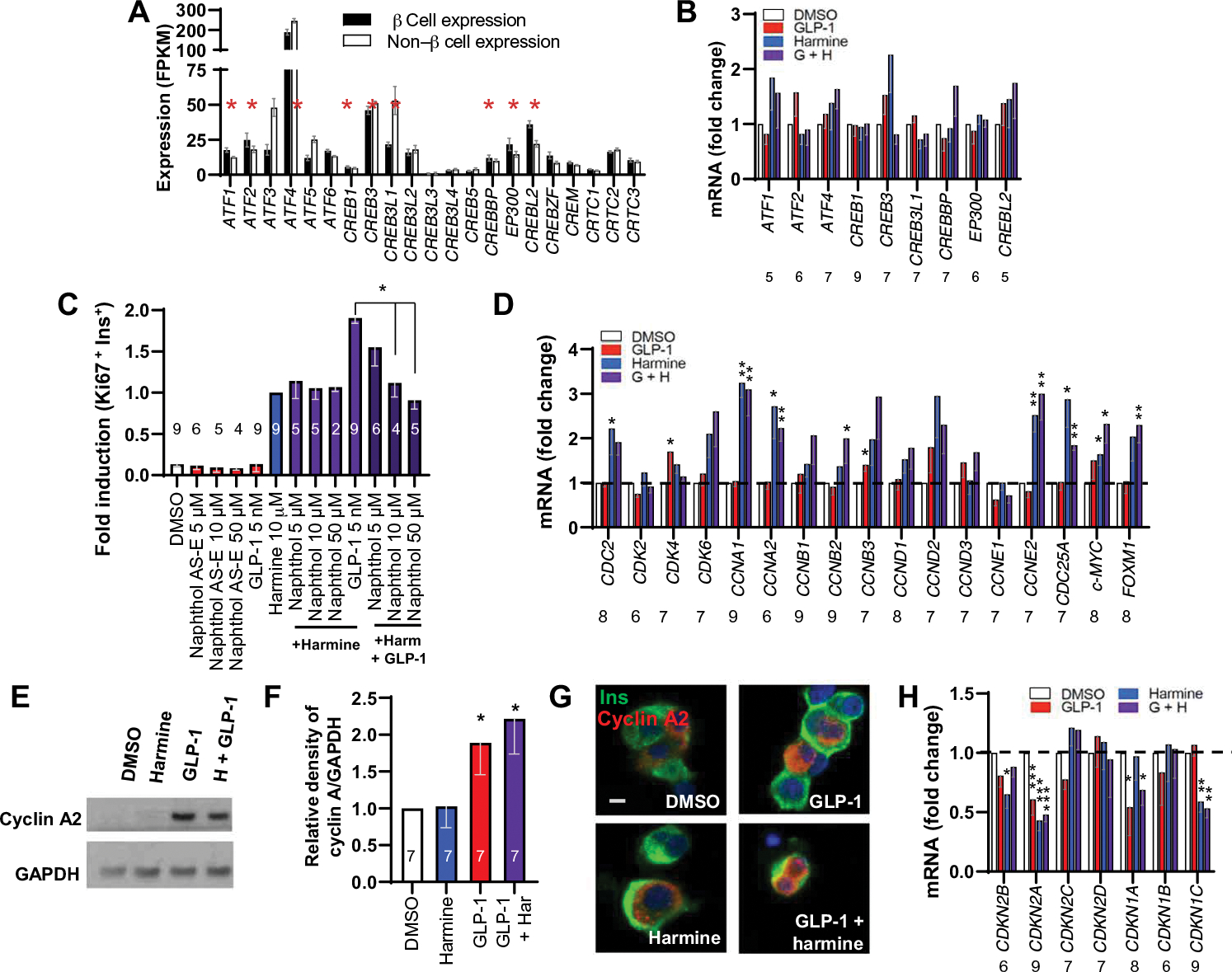
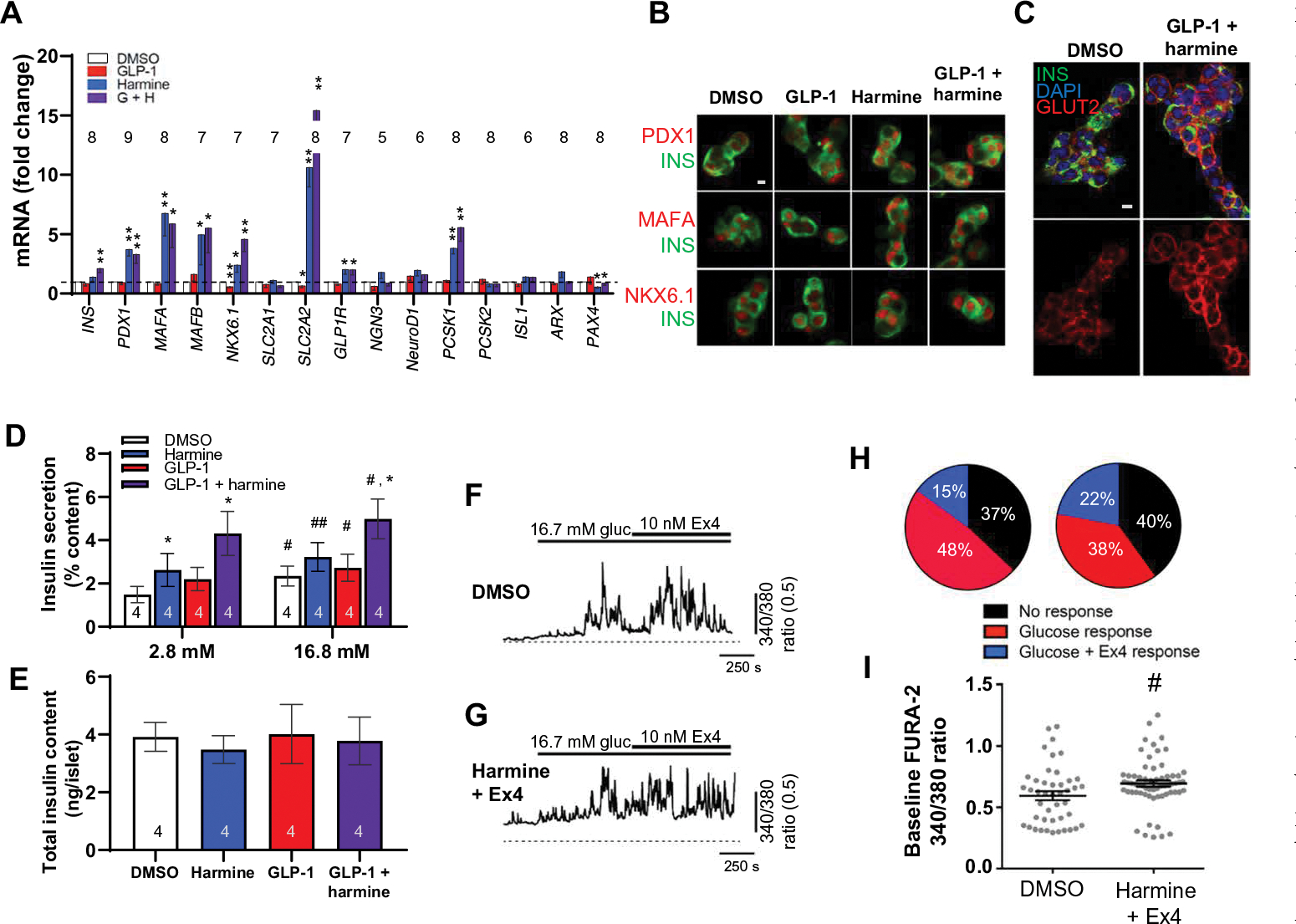
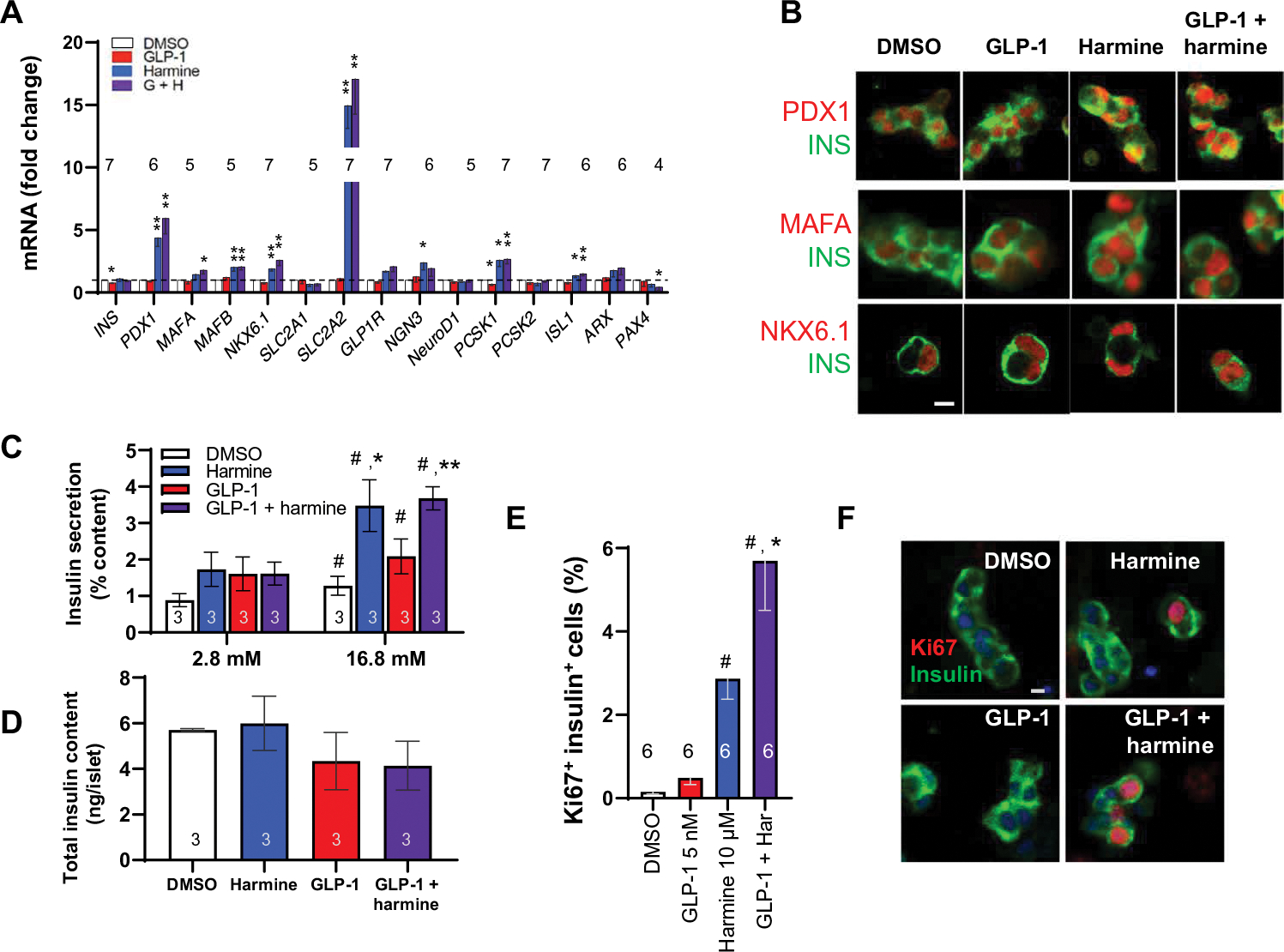
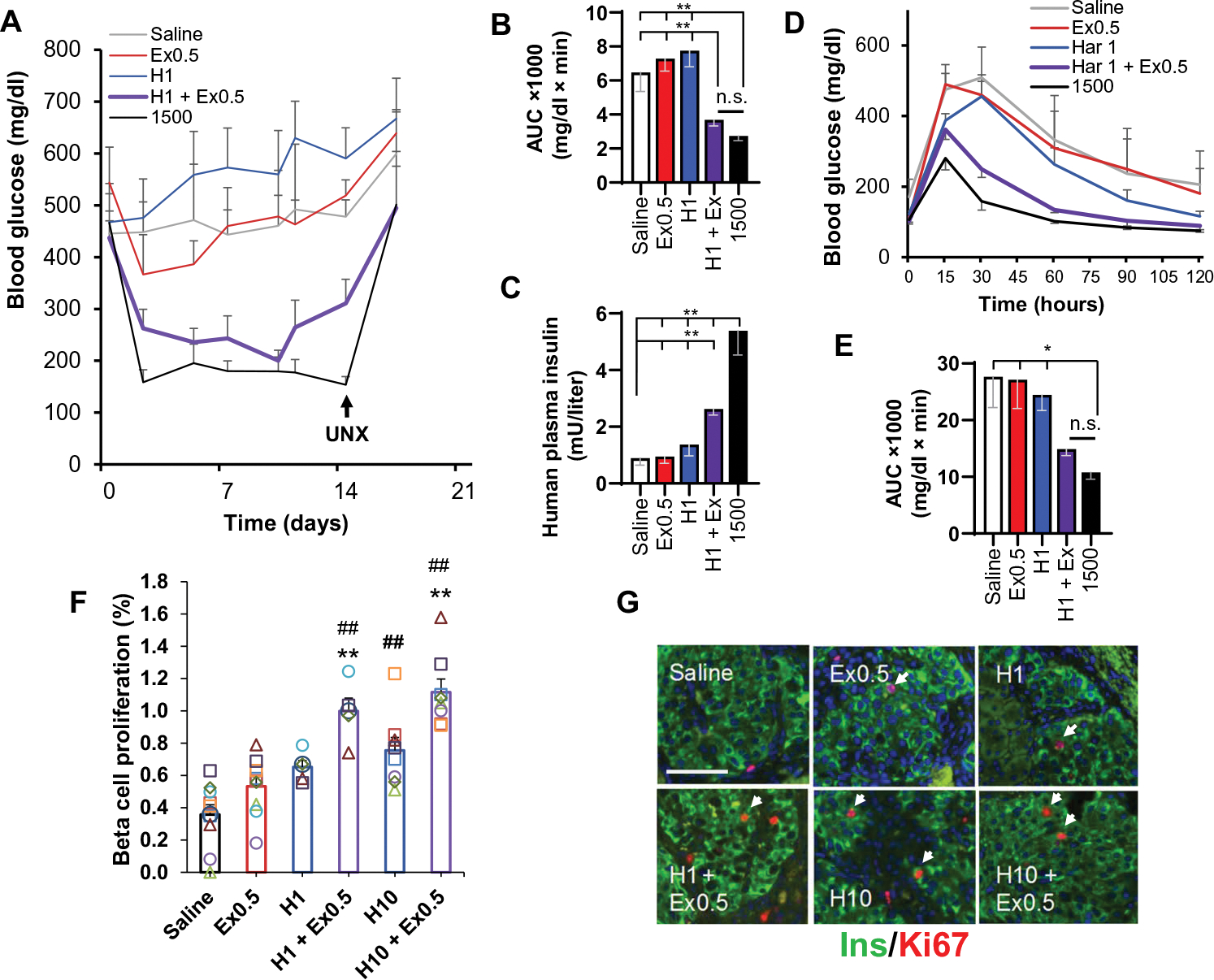
Glucagon-like peptide-1 receptor (GLP1R) agonists and dipeptidyl peptidase 4 inhibitors are widely prescribed diabetes drugs due to their ability to stimulate insulin secretion from remaining β cells and to reduce caloric intake. Unfortunately, they fail to increase human β cell proliferation. Small-molecule inhibitors of dual-specificity tyrosine-regulated kinase 1A (DYRK1A) are able to induce adult human β cell proliferation, but rates are modest (~2%), and their specificity to β cells is limited. Here, we provide evidence that combining any member of the GLP1R agonist class with any member of the DYRK1A inhibitor class induces a synergistic increase in human β cell replication (5 to 6%) accompanied by an actual increase in numbers of human β cells. GLP1R agonist-DYRK1A inhibitor synergy required combined inhibition of DYRK1A and an increase in cAMP and did not lead to β cell dedifferentiation. These beneficial effects on proliferation were seen in both normal human β cells and β cells derived from individuals with type 2 diabetes. The ability of the GLP1R agonist-DYRK1A inhibitor combination to enhance human β cell proliferation, human insulin secretion, and blood glucose control extended in vivo to studies of human islets transplanted into euglycemic and streptozotocin-diabetic immunodeficient mice. No adverse events were observed in the mouse studies during a 1-week period. Because of the relative β cell specificity of GLP1R agonists, the combination provides an improved, although not complete, degree of human β cell specificity.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Two drugs converged in a pancreatic β cell.Sci Transl Med. 2020 Feb 12;12(530):eaba7359. doi: 10.1126/scitranslmed.aba7359. Sci Transl Med. 2020. PMID: 32051228
References
-
- World Health Organization, Global Reports on Diabetes; www.who.int/diabetes/global-report/en/.
-
- Butler AE, Janson J, Bonner-Weir S, Ritzel R, Rizza RA, Butler PC, β-Cell deficit and increased β-cell apoptosis in humans with diabetes. Diabetes 52, 102–110 (2003). - PubMed
-
- Drucker DJ, Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metab. 27, 740–756 (2018). - PubMed
-
- Guo X-H, The value of short- and long-acting glucagon-like peptide agonists in the management of type 2 diabetes mellitus: Experience with exenatide. Curr. Med. Res. Opin. 32, 67–76 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
