

Mutational Functional Analysis of the Pseudorabies Virus Nuclear Egress Complex-Nucleocapsid Interaction
- PMID: 32051272
- PMCID: PMC7108838
- DOI: 10.1128/JVI.01910-19
Mutational Functional Analysis of the Pseudorabies Virus Nuclear Egress Complex-Nucleocapsid Interaction
Abstract
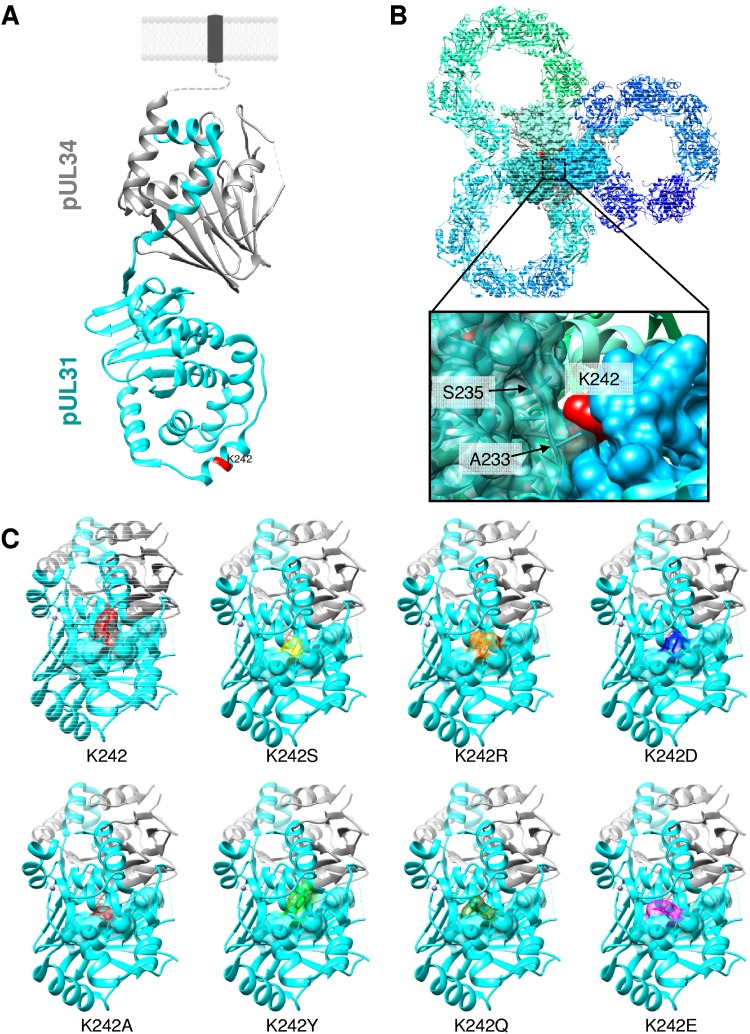
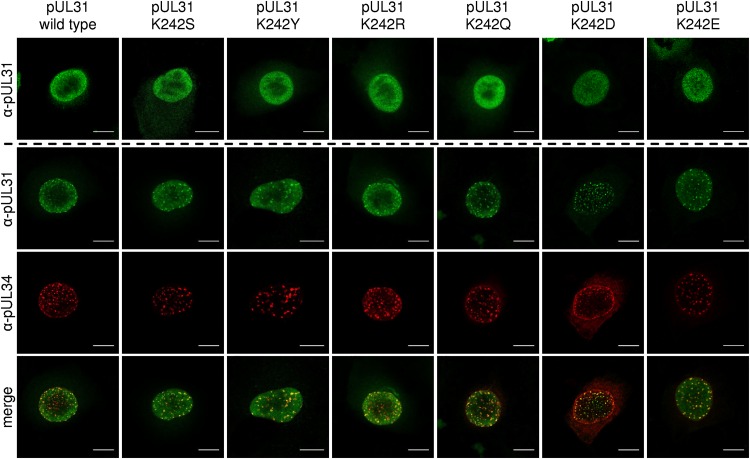
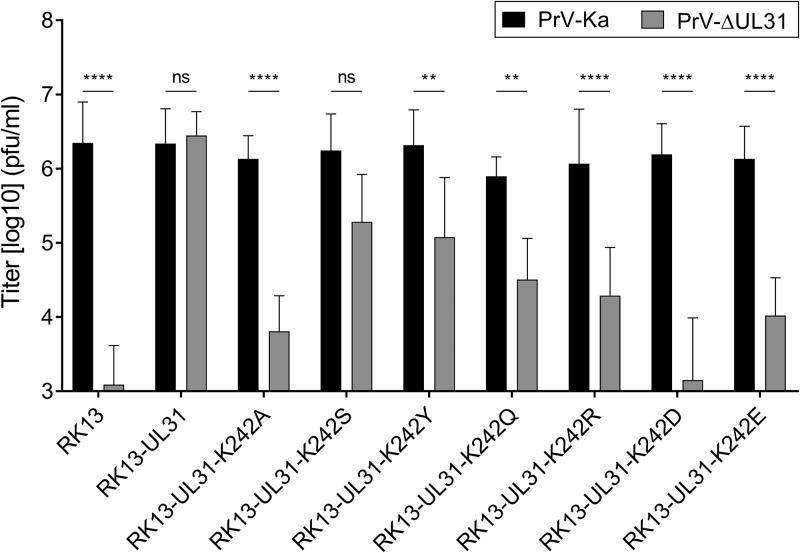
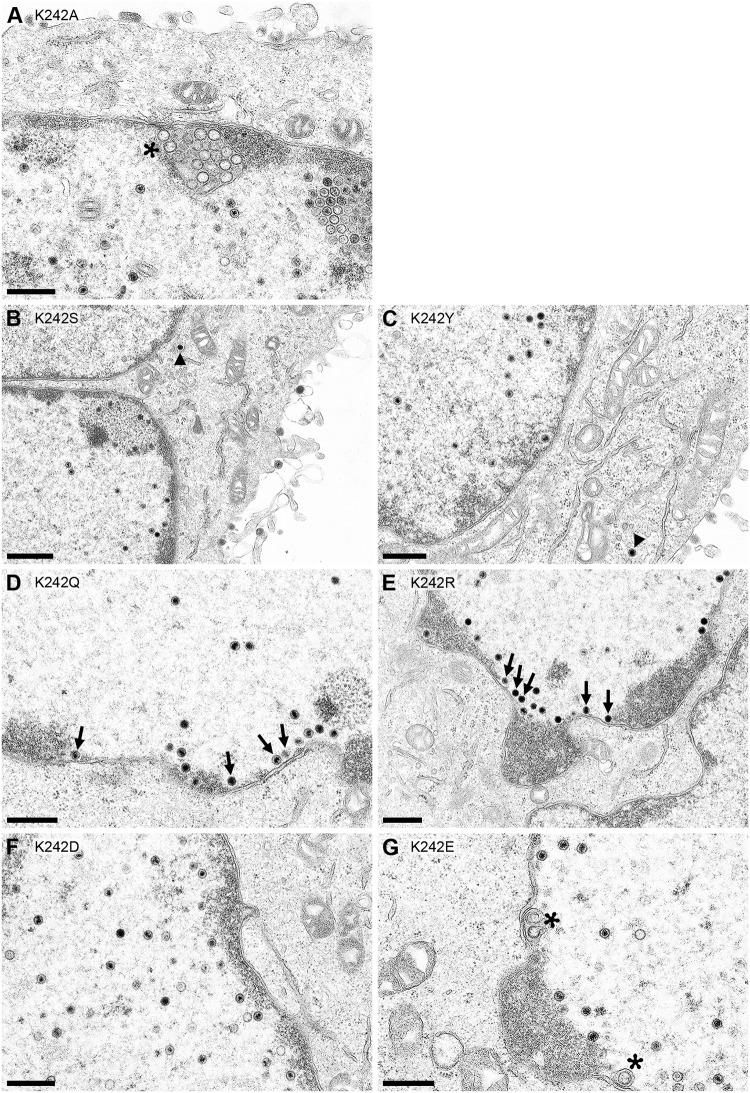
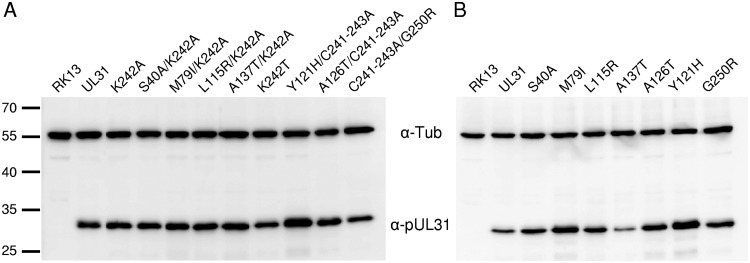
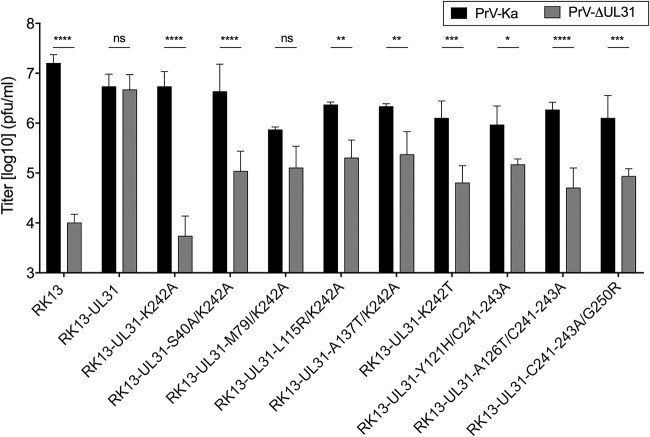
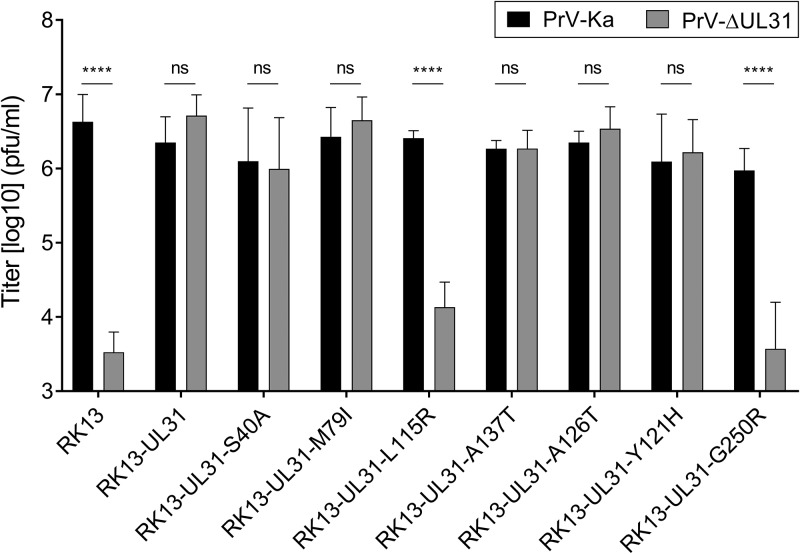
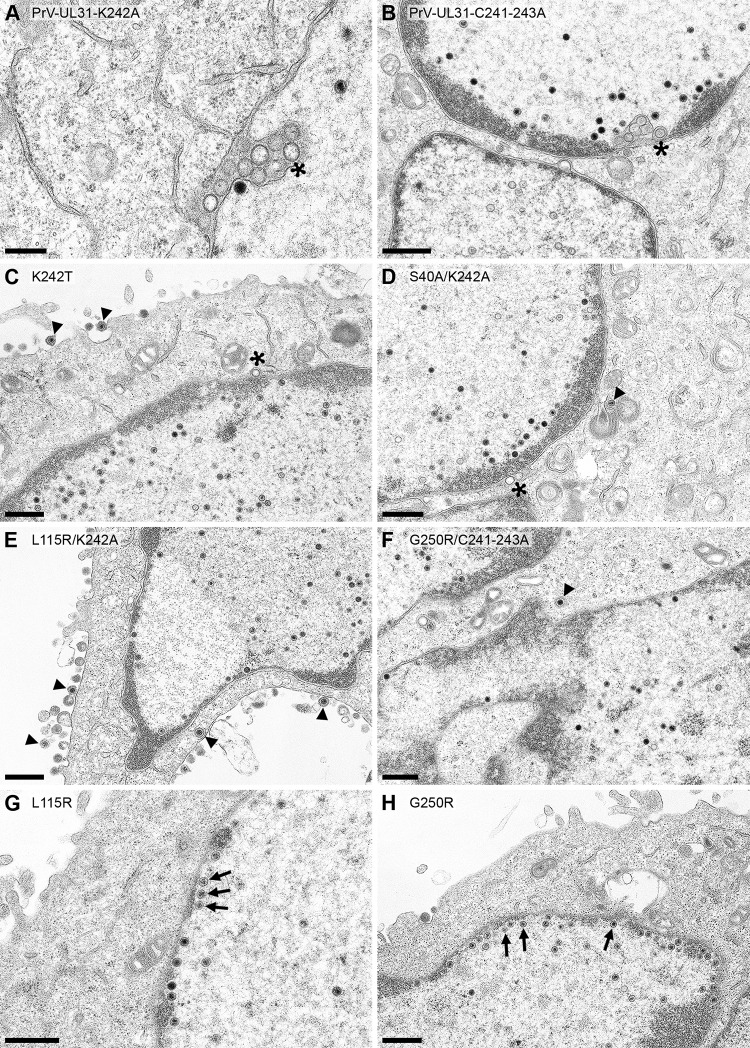
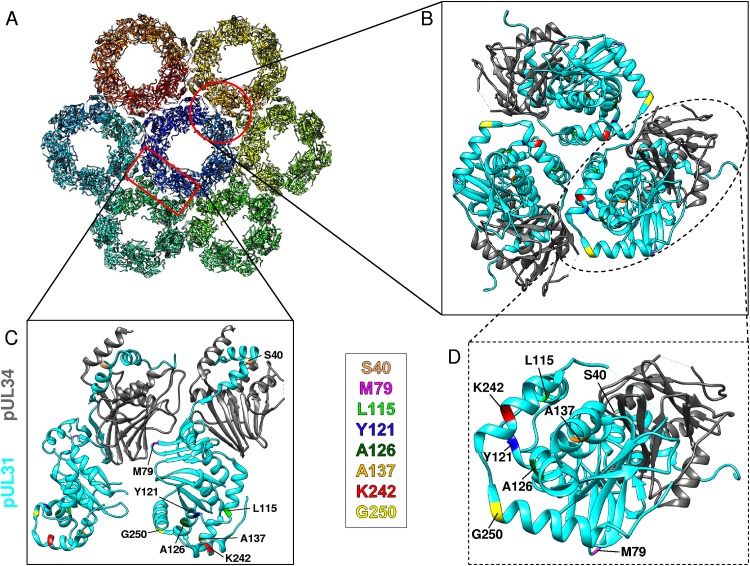
Herpesvirus nucleocapsids leave the nucleus by a vesicle-mediated translocation mediated by the viral nuclear egress complex (NEC). The NEC is composed of two conserved viral proteins, designated pUL34 and pUL31 in the alphaherpesvirus pseudorabies virus (PrV). It is required for efficient nuclear egress and is sufficient for vesicle formation and scission from the inner nuclear membrane (INM). Structure-based mutagenesis identified a lysine at position 242 (K242) in pUL31, located in the most membrane distal part of the NEC, to be crucial for efficient nucleocapsid incorporation into budding vesicles. Replacing the lysine by alanine (K242A) resulted in accumulations of empty vesicles in the perinuclear space, despite the presence of excess nucleocapsids in the nucleus. However, it remained unclear whether the defect in capsid incorporation was due to interference with a direct, electrostatic interaction between the capsid and the NEC or structural restrictions. To test this, we replaced K242 with several amino acids, thereby modifying the charge, size, and side chain orientation. In addition, virus recombinants expressing pUL31-K242A were passaged and screened for second-site mutations. Compensatory mutations at different locations in pUL31 or pUL34 were identified, pointing to an inherent flexibility of the NEC. In summary, our data suggest that the amino acid at position 242 does not directly interact with the nucleocapsid but that rearrangements in the NEC coat are required for efficient nucleocapsid envelopment at the INM.IMPORTANCE Herpesviruses encode an exceptional vesicle formation and scission machinery, which operates at the inner nuclear membrane, translocating the viral nucleocapsid from the nucleus into the perinuclear space. The conserved herpesviral nuclear egress complex (NEC) orchestrates this process. High-resolution imaging approaches as well as the recently solved crystal structures of the NEC provided deep insight into the molecular details of vesicle formation and scission. Nevertheless, the molecular mechanism of nucleocapsid incorporation remained unclear. In accordance with structure-based predictions, a basic amino acid could be pinpointed in the most membrane-distal domain of the NEC (pUL31-K242), indicating that capsid incorporation might depend on a direct electrostatic interaction. Our follow-up study, described here, however, shows that the positive charge is not relevant but that the overall structure matters.
Keywords: herpesvirus; nuclear egress complex; nuclear envelope; pUL31; pUL34; pseudorabies virus.
Copyright © 2020 American Society for Microbiology.
Figures
Similar articles
-
Lysine 242 within Helix 10 of the Pseudorabies Virus Nuclear Egress Complex pUL31 Component Is Critical for Primary Envelopment of Nucleocapsids.J Virol. 2017 Oct 27;91(22):e01182-17. doi: 10.1128/JVI.01182-17. Print 2017 Nov 15. J Virol. 2017. PMID: 28878082 Free PMC article.
-
Function of the Nonconserved N-Terminal Domain of Pseudorabies Virus pUL31 in Nuclear Egress.J Virol. 2018 Jul 17;92(15):e00566-18. doi: 10.1128/JVI.00566-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29793954 Free PMC article.
-
Generation and characterization of monoclonal antibodies specific for the Pseudorabies Virus nuclear egress complex.Virus Res. 2020 Oct 2;287:198096. doi: 10.1016/j.virusres.2020.198096. Epub 2020 Jul 17. Virus Res. 2020. PMID: 32682818
-
Venture from the Interior-Herpesvirus pUL31 Escorts Capsids from Nucleoplasmic Replication Compartments to Sites of Primary Envelopment at the Inner Nuclear Membrane.Cells. 2017 Nov 25;6(4):46. doi: 10.3390/cells6040046. Cells. 2017. PMID: 29186822 Free PMC article. Review.
-
The Knowns and Unknowns of Herpesvirus Nuclear Egress.Annu Rev Virol. 2023 Sep 29;10(1):305-323. doi: 10.1146/annurev-virology-111821-105518. Epub 2023 Apr 11. Annu Rev Virol. 2023. PMID: 37040797 Review.
Cited by
-
Nuclear Egress.Curr Issues Mol Biol. 2021;41:125-170. doi: 10.21775/cimb.041.125. Epub 2020 Aug 7. Curr Issues Mol Biol. 2021. PMID: 32764158 Free PMC article. Review.
-
Alphaherpesvirus in Pets and Livestock.Microorganisms. 2025 Jan 4;13(1):82. doi: 10.3390/microorganisms13010082. Microorganisms. 2025. PMID: 39858850 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
