Periodic Parasites and Daily Host Rhythms
- PMID: 32053788
- PMCID: PMC7137616
- DOI: 10.1016/j.chom.2020.01.005
Periodic Parasites and Daily Host Rhythms
Abstract
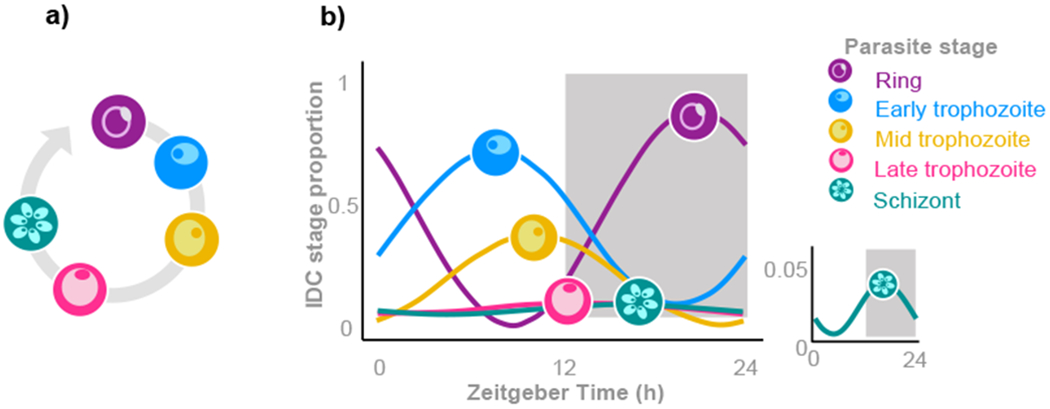
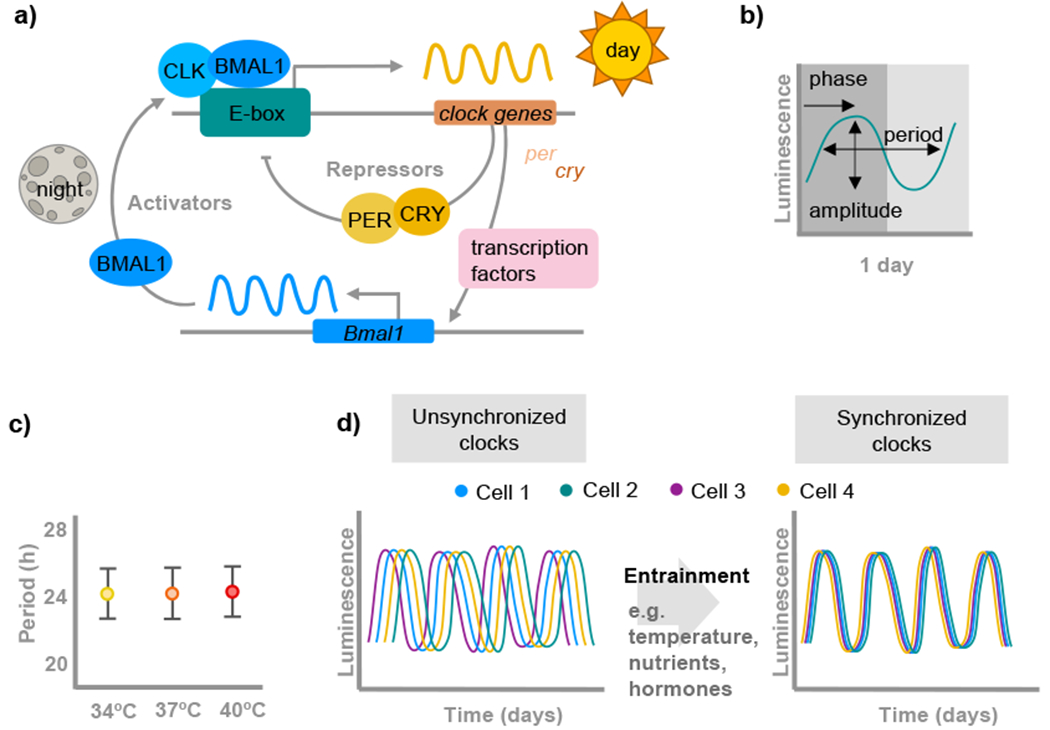
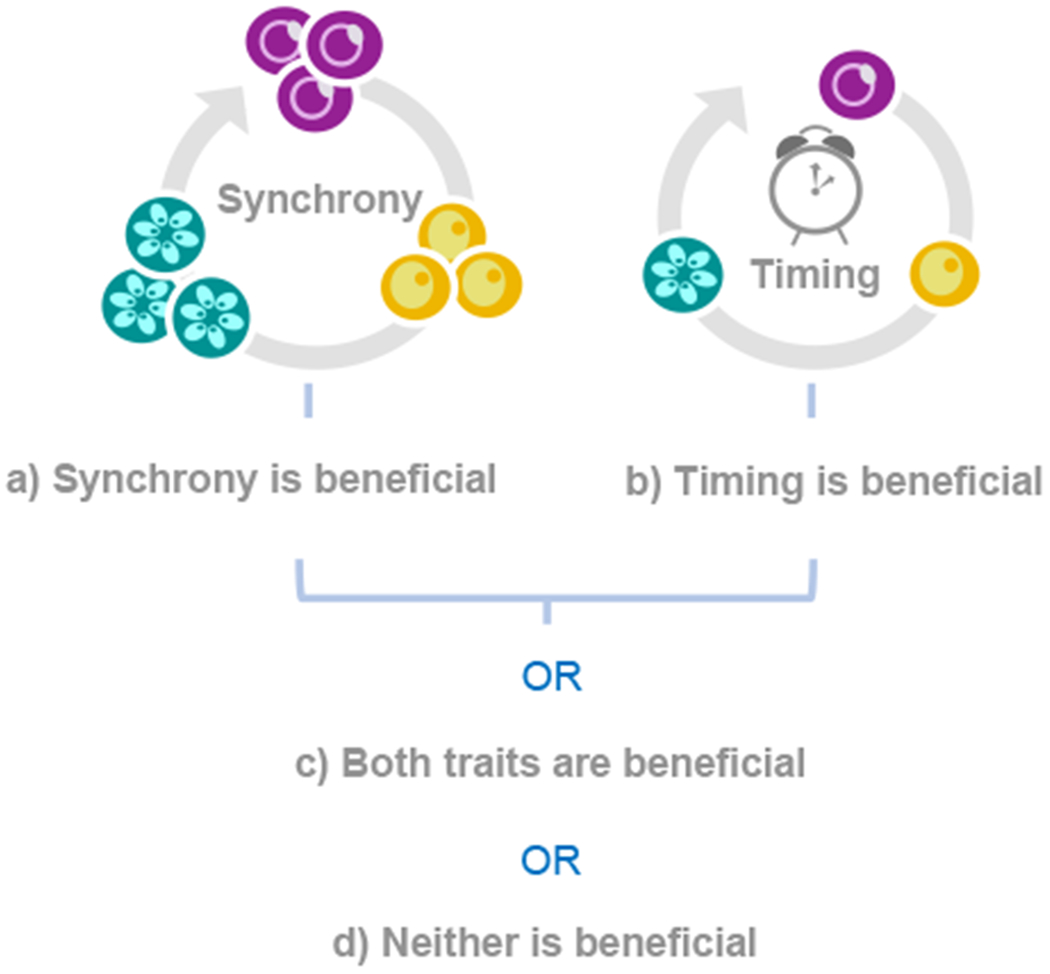
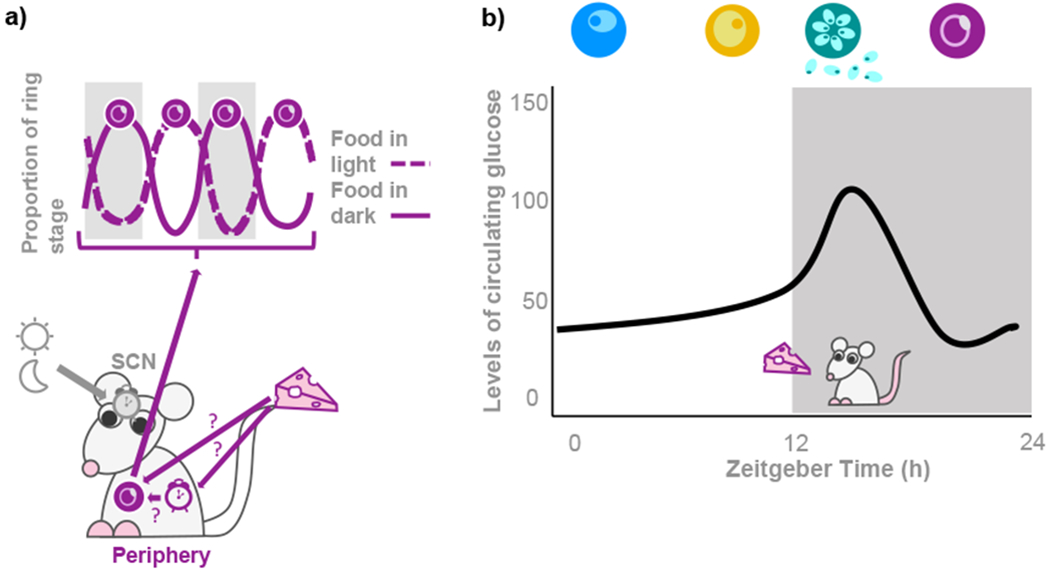
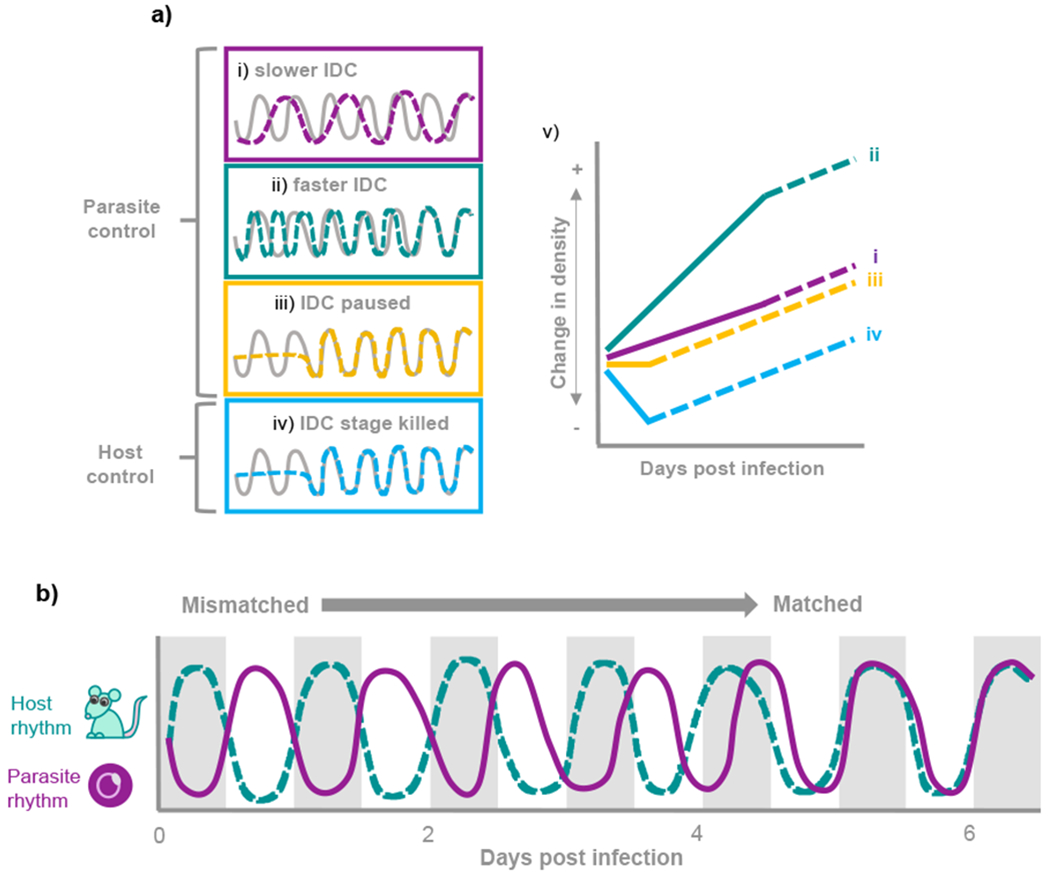
Biological rhythms appear to be an elegant solution to the challenge of coordinating activities with the consequences of the Earth's daily and seasonal rotation. The genes and molecular mechanisms underpinning circadian clocks in multicellular organisms are well understood. In contrast, the regulatory mechanisms and fitness consequences of biological rhythms exhibited by parasites remain mysterious. Here, we explore how periodicity in parasite traits is generated and why daily rhythms matter for parasite fitness. We focus on malaria (Plasmodium) parasites which exhibit developmental rhythms during replication in the mammalian host's blood and in transmission to vectors. Rhythmic in-host parasite replication is responsible for eliciting inflammatory responses, the severity of disease symptoms, and fueling transmission, as well as conferring tolerance to anti-parasite drugs. Thus, understanding both how and why the timing and synchrony of parasites are connected to the daily rhythms of hosts and vectors may make treatment more effective and less toxic to hosts.
Keywords: Plasmodium; circadian clock; circadian rhythm; entrainment; fitness; host-parasite interactions; inflammatory response; intra-erythrocytic development cycle; metabolism; nutrient sensing; periodicity; synchronicity.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures