

Reduced Function of the Glutathione S-Transferase S1 Suppresses Behavioral Hyperexcitability in Drosophila Expressing Mutant Voltage-Gated Sodium Channels
- PMID: 32054635
- PMCID: PMC7144092
- DOI: 10.1534/g3.119.401025
Reduced Function of the Glutathione S-Transferase S1 Suppresses Behavioral Hyperexcitability in Drosophila Expressing Mutant Voltage-Gated Sodium Channels
Abstract
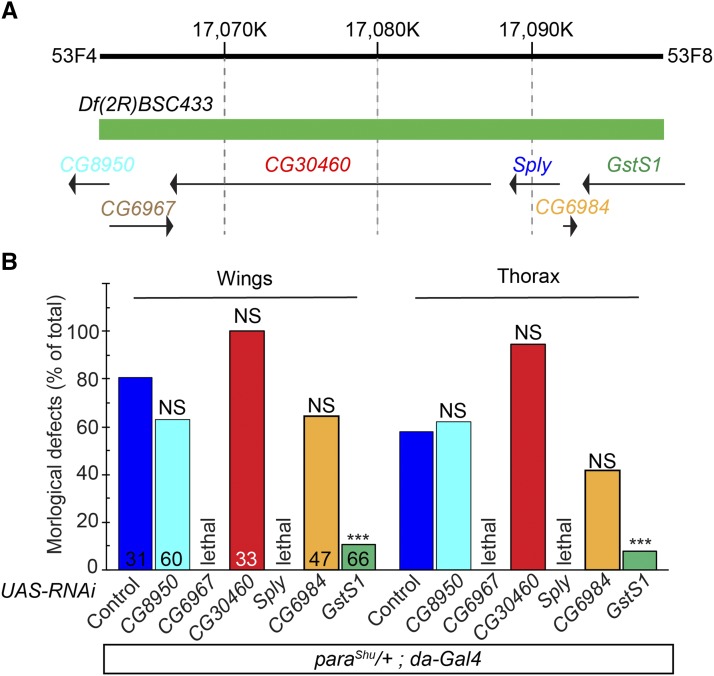
Voltage-gated sodium (Nav) channels play a central role in the generation and propagation of action potentials in excitable cells such as neurons and muscles. To determine how the phenotypes of Nav-channel mutants are affected by other genes, we performed a forward genetic screen for dominant modifiers of the seizure-prone, gain-of-function Drosophila melanogaster Nav-channel mutant, paraShu Our analyses using chromosome deficiencies, gene-specific RNA interference, and single-gene mutants revealed that a null allele of glutathione S-transferase S1 (GstS1) dominantly suppresses paraShu phenotypes. Reduced GstS1 function also suppressed phenotypes of other seizure-prone Nav-channel mutants, paraGEFS+ and parabss Notably, paraShu mutants expressed 50% less GstS1 than wild-type flies, further supporting the notion that paraShu and GstS1 interact functionally. Introduction of a loss-of-function GstS1 mutation into a paraShu background led to up- and down-regulation of various genes, with those encoding cytochrome P450 (CYP) enzymes most significantly over-represented in this group. Because GstS1 is a fly ortholog of mammalian hematopoietic prostaglandin D synthase, and in mammals CYPs are involved in the oxygenation of polyunsaturated fatty acids including prostaglandins, our results raise the intriguing possibility that bioactive lipids play a role in GstS1-mediated suppression of paraShu phenotypes.
Keywords: Forward genetic screen; RNA-sequencing analysis; epilepsy; genetic modifiers.
Copyright © 2020 Chen et al.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases