

The antiarrhythmic compound efsevin directly modulates voltage-dependent anion channel 2 by binding to its inner wall and enhancing mitochondrial Ca2+ uptake
- PMID: 32059260
- PMCID: PMC7279994
- DOI: 10.1111/bph.15022
The antiarrhythmic compound efsevin directly modulates voltage-dependent anion channel 2 by binding to its inner wall and enhancing mitochondrial Ca2+ uptake
Abstract
Background and purpose: The synthetic compound efsevin was recently identified to suppress arrhythmogenesis in models of cardiac arrhythmia, making it a promising candidate for antiarrhythmic therapy. Its activity was shown to be dependent on the voltage-dependent anion channel 2 (VDAC2) in the outer mitochondrial membrane. Here, we investigated the molecular mechanism of the efsevin-VDAC2 interaction.
Experimental approach: To evaluate the functional interaction of efsevin and VDAC2, we measured currents through recombinant VDAC2 in planar lipid bilayers. Using molecular ligand-protein docking and mutational analysis, we identified the efsevin binding site on VDAC2. Finally, physiological consequences of the efsevin-induced modulation of VDAC2 were analysed in HL-1 cardiomyocytes.
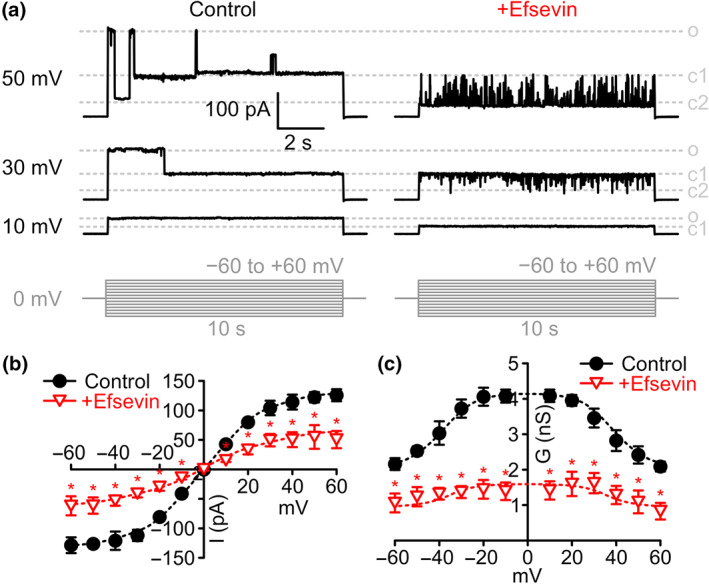
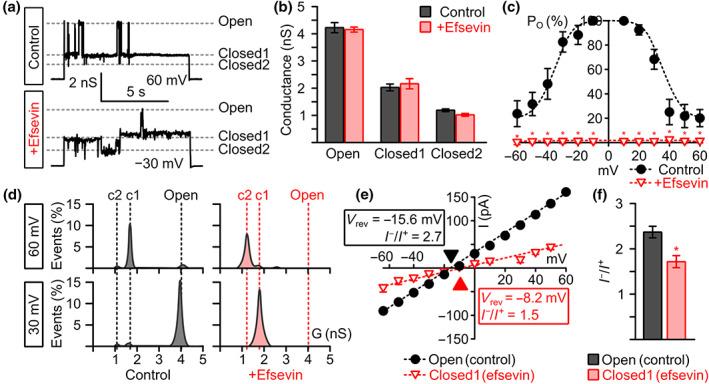
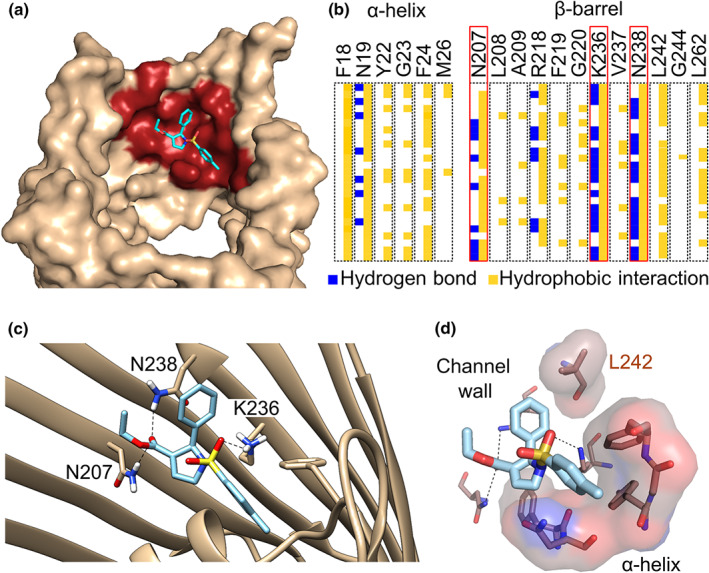
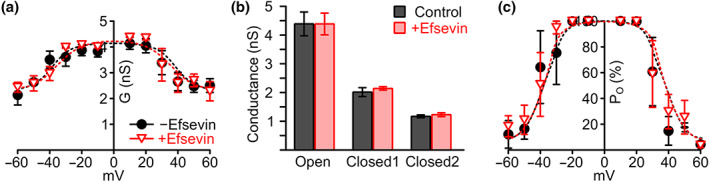
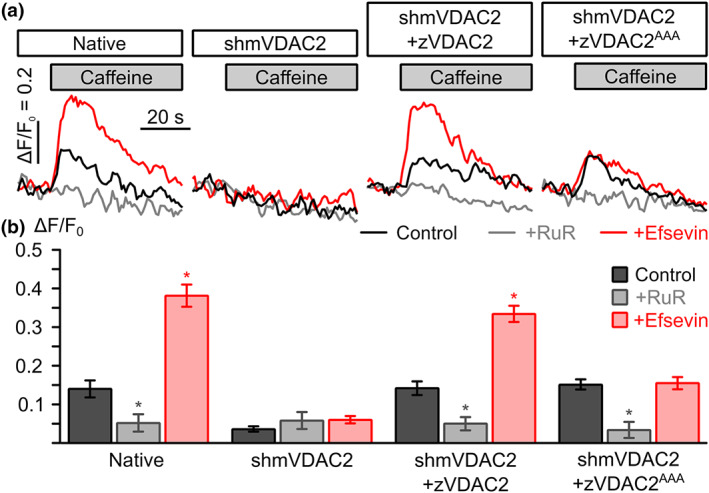
Key results: In lipid bilayers, efsevin reduced VDAC2 conductance and shifted the channel's open probability towards less anion-selective closed states. Efsevin binds to a binding pocket formed by the inner channel wall and the pore-lining N-terminal α-helix. Exchange of amino acids N207, K236 and N238 within this pocket for alanines abolished the channel's efsevin-responsiveness. Upon heterologous expression in HL-1 cardiomyocytes, both channels, wild-type VDAC2 and the efsevin-insensitive VDAC2AAA restored mitochondrial Ca2+ uptake, but only wild-type VDAC2 was sensitive to efsevin.
Conclusion and implications: In summary, our data indicate a direct interaction of efsevin with VDAC2 inside the channel pore that leads to modified gating and results in enhanced SR-mitochondria Ca2+ transfer. This study sheds new light on the function of VDAC2 and provides a basis for structure-aided chemical optimization of efsevin.
© 2020 The Authors. British Journal of Pharmacology published by John Wiley & Sons Ltd on behalf of British Pharmacological Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Mitochondrial Ca(2+) uptake by the voltage-dependent anion channel 2 regulates cardiac rhythmicity.Elife. 2015 Jan 15;4:e04801. doi: 10.7554/eLife.04801. Elife. 2015. PMID: 25588501 Free PMC article.
-
Cardiac-specific deletion of voltage dependent anion channel 2 leads to dilated cardiomyopathy by altering calcium homeostasis.Nat Commun. 2021 Jul 28;12(1):4583. doi: 10.1038/s41467-021-24869-0. Nat Commun. 2021. PMID: 34321484 Free PMC article.
-
Voltage-dependent anion channel 2 modulates resting Ca²+ sparks, but not action potential-induced Ca²+ signaling in cardiac myocytes.Cell Calcium. 2011 Feb;49(2):136-43. doi: 10.1016/j.ceca.2010.12.004. Epub 2011 Jan 15. Cell Calcium. 2011. PMID: 21241999
-
VDAC2 as a novel target for heart failure: Ca2+ at the sarcomere, mitochondria and SR.Cell Calcium. 2022 Jun;104:102586. doi: 10.1016/j.ceca.2022.102586. Epub 2022 Mar 28. Cell Calcium. 2022. PMID: 35429733 Free PMC article. Review.
-
VDAC2-specific cellular functions and the underlying structure.Biochim Biophys Acta. 2016 Oct;1863(10):2503-14. doi: 10.1016/j.bbamcr.2016.04.020. Epub 2016 Apr 23. Biochim Biophys Acta. 2016. PMID: 27116927 Free PMC article. Review.
Cited by
-
Cellular and mitochondrial mechanisms of atrial fibrillation.Basic Res Cardiol. 2020 Nov 30;115(6):72. doi: 10.1007/s00395-020-00827-7. Basic Res Cardiol. 2020. PMID: 33258071 Free PMC article. Review.
-
E as in Enigma: The Mysterious Role of the Voltage-Dependent Anion Channel Glutamate E73.Int J Mol Sci. 2022 Dec 23;24(1):269. doi: 10.3390/ijms24010269. Int J Mol Sci. 2022. PMID: 36613710 Free PMC article. Review.
-
STING Suppresses Mitochondrial VDAC2 to Govern RCC Growth Independent of Innate Immunity.Adv Sci (Weinh). 2023 Jan;10(3):e2203718. doi: 10.1002/advs.202203718. Epub 2022 Nov 29. Adv Sci (Weinh). 2023. PMID: 36445063 Free PMC article.
-
Therapeutic Strategies Targeting Mitochondrial Calcium Signaling: A New Hope for Neurological Diseases?Antioxidants (Basel). 2022 Jan 15;11(1):165. doi: 10.3390/antiox11010165. Antioxidants (Basel). 2022. PMID: 35052668 Free PMC article. Review.
-
Metabolic remodelling in atrial fibrillation: manifestations, mechanisms and clinical implications.Nat Rev Cardiol. 2024 Oct;21(10):682-700. doi: 10.1038/s41569-024-01038-6. Epub 2024 May 30. Nat Rev Cardiol. 2024. PMID: 38816507 Review.
References
-
- Benjamin, E. J. , Virani, S. S. , Callaway, C. W. , Chamberlain, A. M. , Chang, A. R. , Cheng, S. , … American Heart Association Council on Epidemiology and Prevention Statistics Committee and Stroke Statistics Subcommittee . (2018). Heart disease and stroke statistics—2018 update: A report from the American Heart Association. Circulation, 137, e67–e492. 10.1161/CIR.0000000000000558 - DOI - PubMed
-
- Claycomb, W. C. , Lanson, N. A. , Stallworth, B. S. , Egeland, D. B. , Delcarpio, J. B. , Bahinski, A. , & Izzo, N. J. (1998). HL‐1 cells: A cardiac muscle cell line that contracts and retains phenotypic characteristics of the adult cardiomyocyte. Proceedings of the National Academy of Sciences of the United States of America, 95, 2979–2984. 10.1073/pnas.95.6.2979 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
