

Biosurfactant-Mediated Membrane Depolarization Maintains Viability during Oxygen Depletion in Bacillus subtilis
- PMID: 32059765
- PMCID: PMC7153240
- DOI: 10.1016/j.cub.2020.01.073
Biosurfactant-Mediated Membrane Depolarization Maintains Viability during Oxygen Depletion in Bacillus subtilis
Abstract
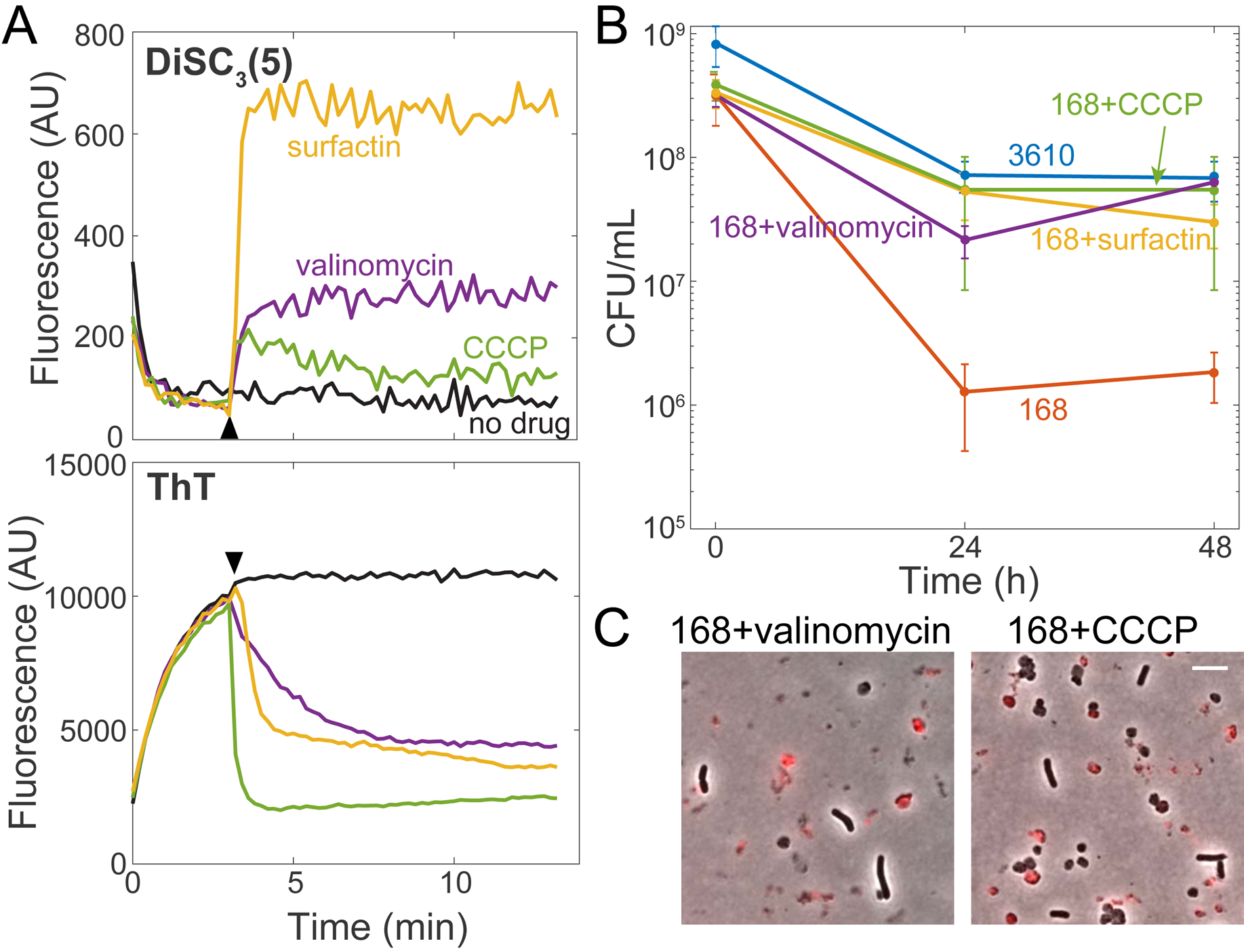
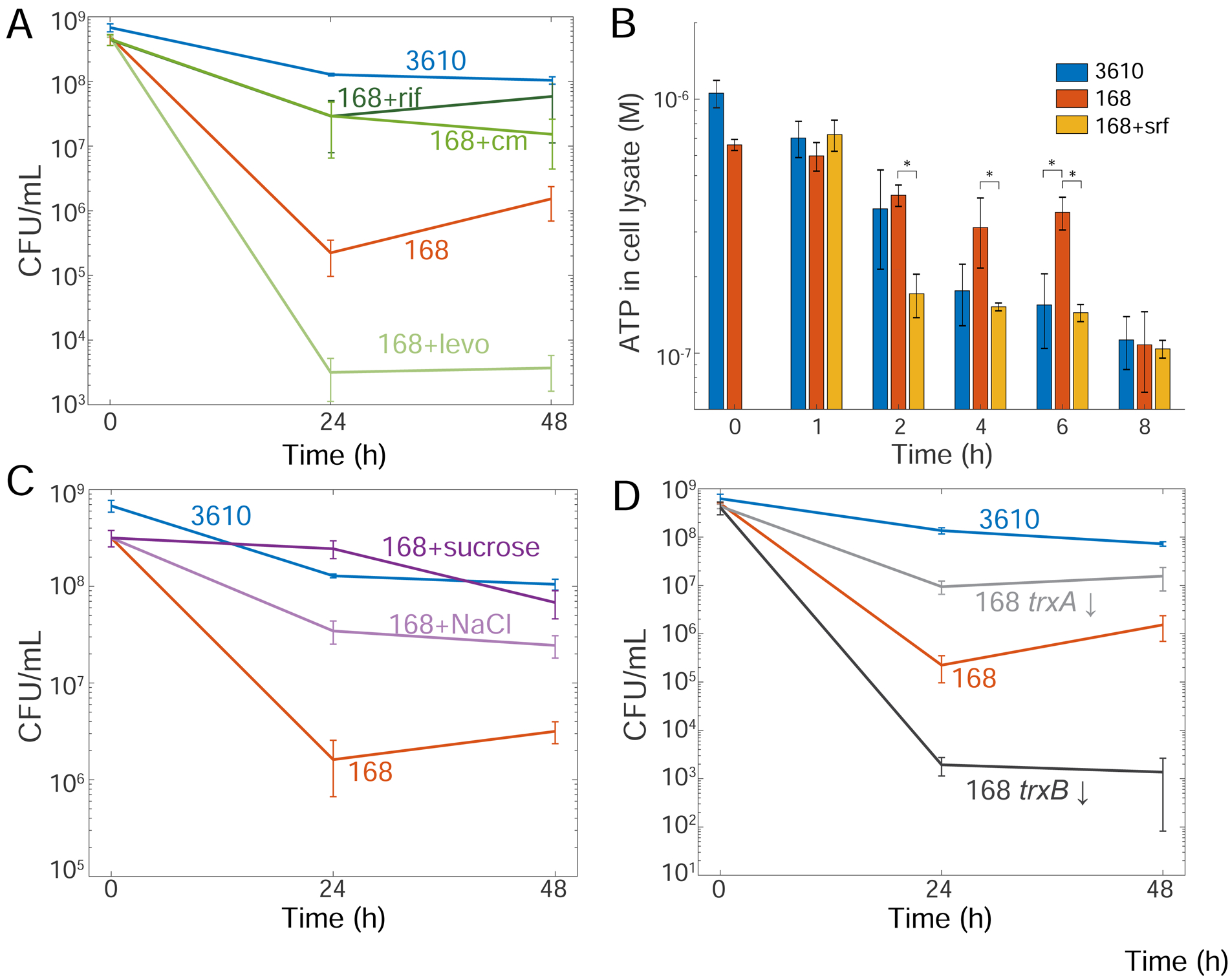
The presence or absence of oxygen in the environment is a strong effector of cellular metabolism and physiology. Like many eukaryotes and some bacteria, Bacillus subtilis primarily utilizes oxygen during respiration to generate ATP. Despite the importance of oxygen for B. subtilis survival, we know little about how populations adapt to shifts in oxygen availability. Here, we find that when oxygen was depleted from stationary phase B. subtilis cultures, ∼90% of cells died while the remaining cells maintained colony-forming ability. We discover that production of the antimicrobial surfactin confers two oxygen-related fitness benefits: it increases aerobic growth yield by increasing oxygen diffusion, and it maintains viability during oxygen depletion by depolarizing the membrane. Strains unable to produce surfactin exhibited an ∼50-fold reduction in viability after oxygen depletion. Surfactin treatment of these cells led to membrane depolarization and reduced ATP production. Chemical and genetic perturbations that alter oxygen consumption or redox state support a model in which surfactin-mediated membrane depolarization maintains viability through slower oxygen consumption and/or a shift to a more reduced metabolic profile. These findings highlight the importance of membrane potential in regulating cell physiology and growth, and demonstrate that antimicrobials that depolarize cell membranes can benefit cells when the terminal electron acceptor in respiration is limiting. This foundational knowledge has deep implications for environmental microbiology, clinical anti-bacterial therapy, and industrial biotechnology.
Keywords: aerobic respiration; biosurfactant; cell lysis; hypoxia; membrane depolarization; membrane potential; oxygen depletion; oxygen diffusion; surfactin.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
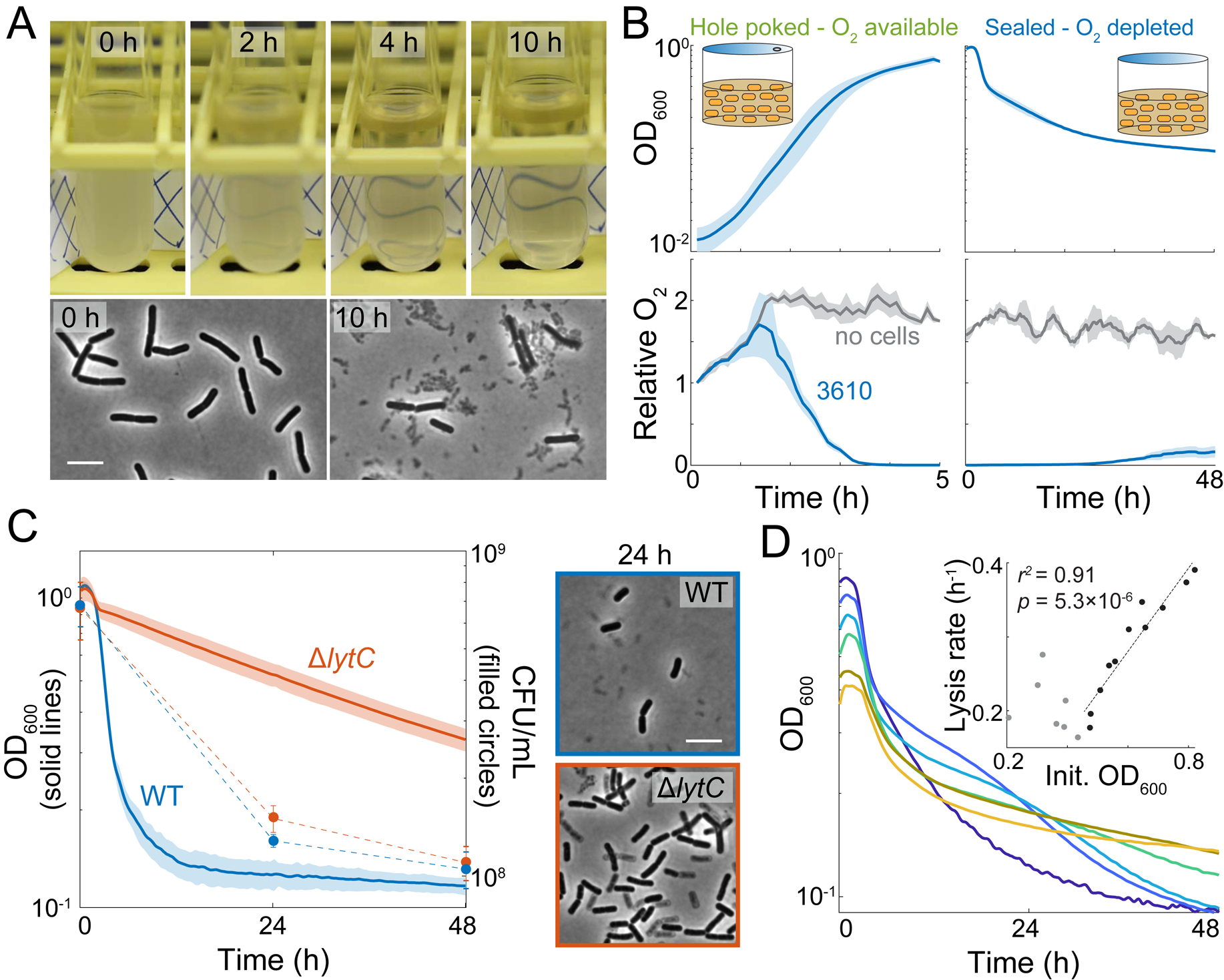
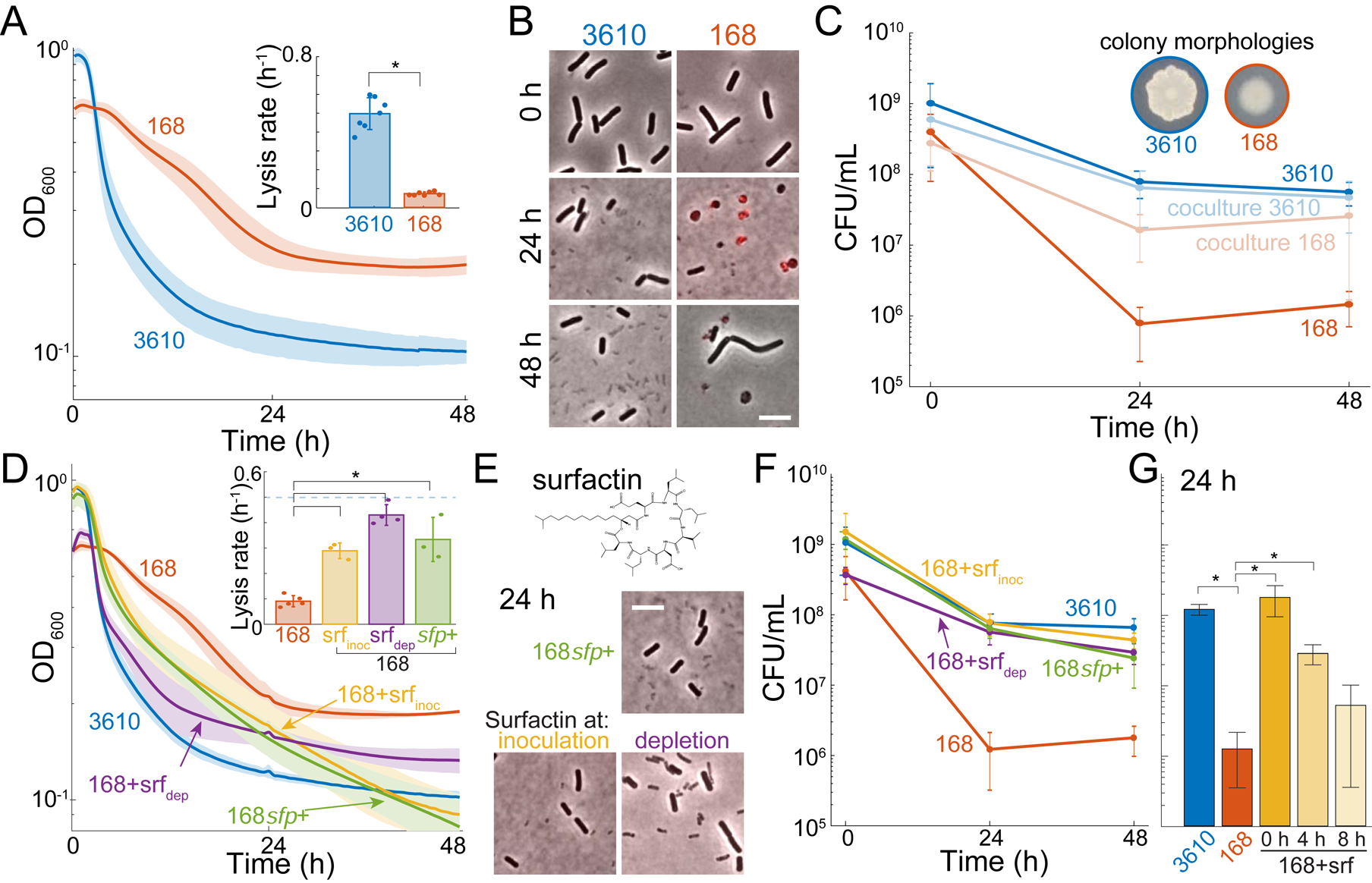
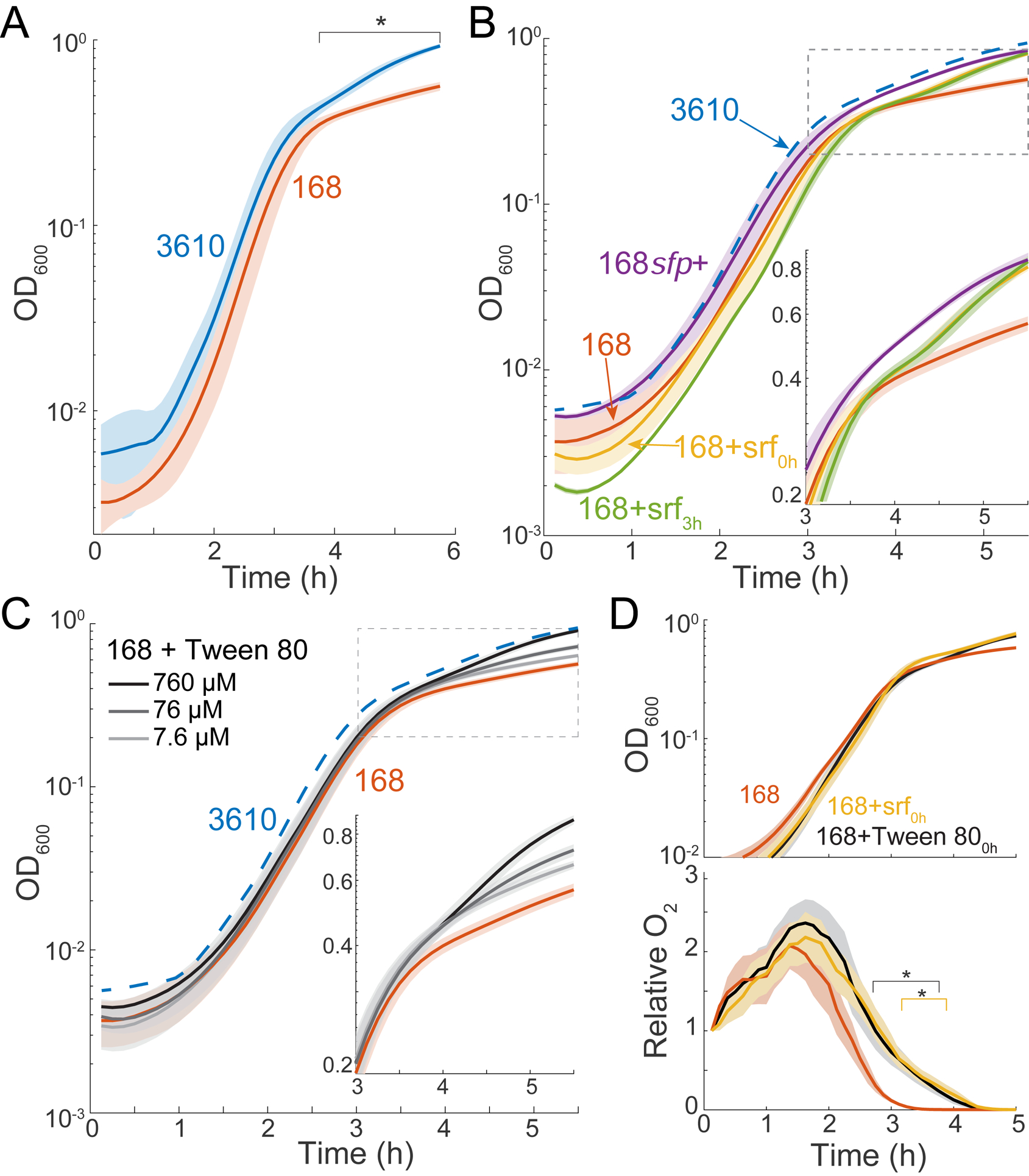
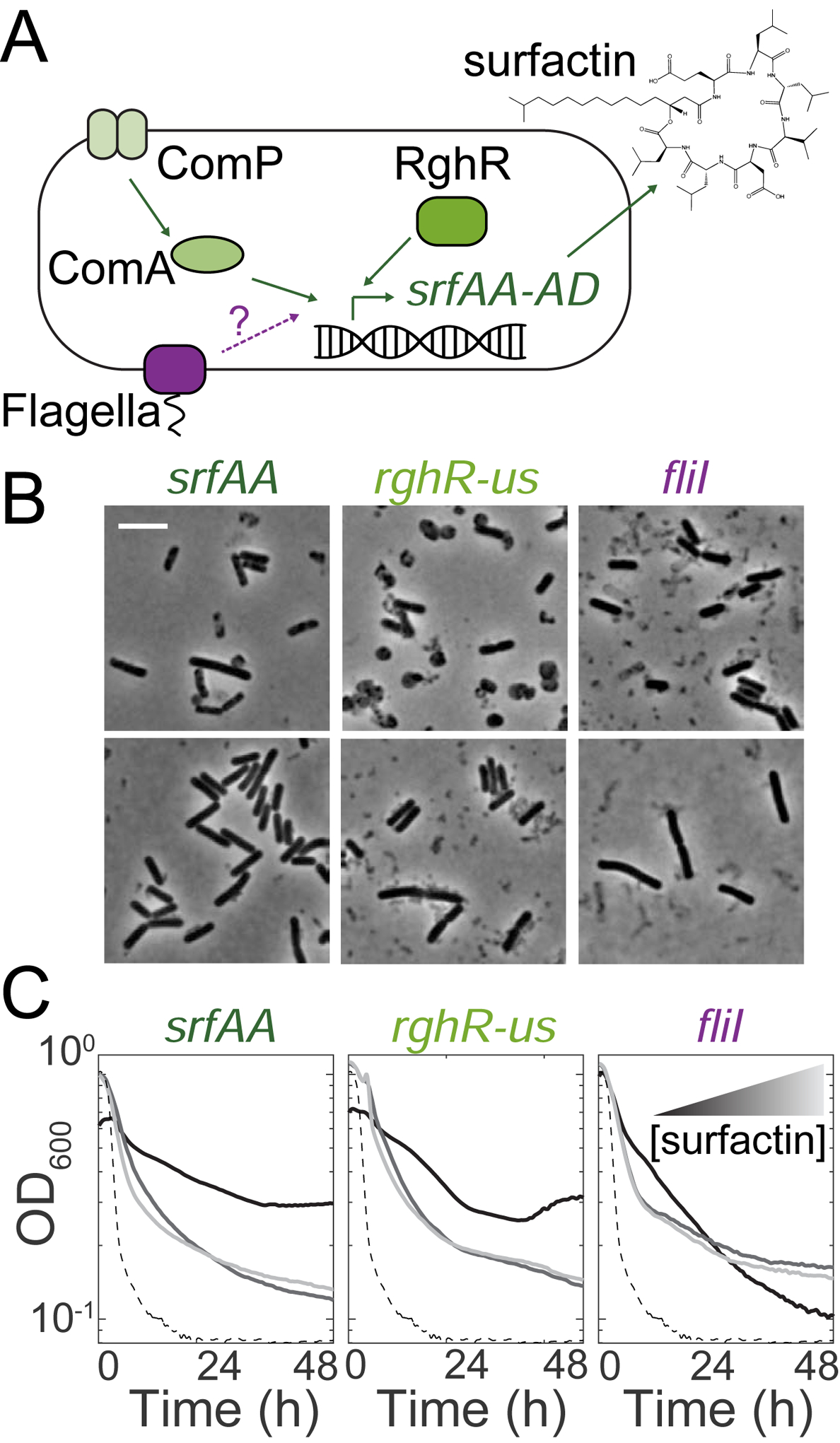
Comment in
-
Anaerobiosis: A Surfactant Helps Bacteria Breathe a Sigh of Relief.Curr Biol. 2020 Mar 23;30(6):R278-R280. doi: 10.1016/j.cub.2020.01.087. Curr Biol. 2020. PMID: 32208155
References
-
- Boutilier RG (2001). Mechanisms of cell survival in hypoxia and hypothermia. J Exp Biol 204, 3171–3181. - PubMed
-
- Hochachka PW (1986). Defense strategies against hypoxia and hypothermia. Science 231, 234–241. - PubMed
-
- Sendoel A, and Hengartner MO (2014). Apoptotic cell death under hypoxia. Physiology (Bethesda) 29, 168–176. - PubMed
-
- Prescott LM, Harley JP, and Klein DA (2005). Microbiology, 6th Edition, (Dubuque, IA: McGraw-Hill Higher Education; ).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
