

GROWTH-REGULATING FACTORS Interact with DELLAs and Regulate Growth in Cold Stress
- PMID: 32060178
- PMCID: PMC7145461
- DOI: 10.1105/tpc.19.00784
GROWTH-REGULATING FACTORS Interact with DELLAs and Regulate Growth in Cold Stress
Abstract
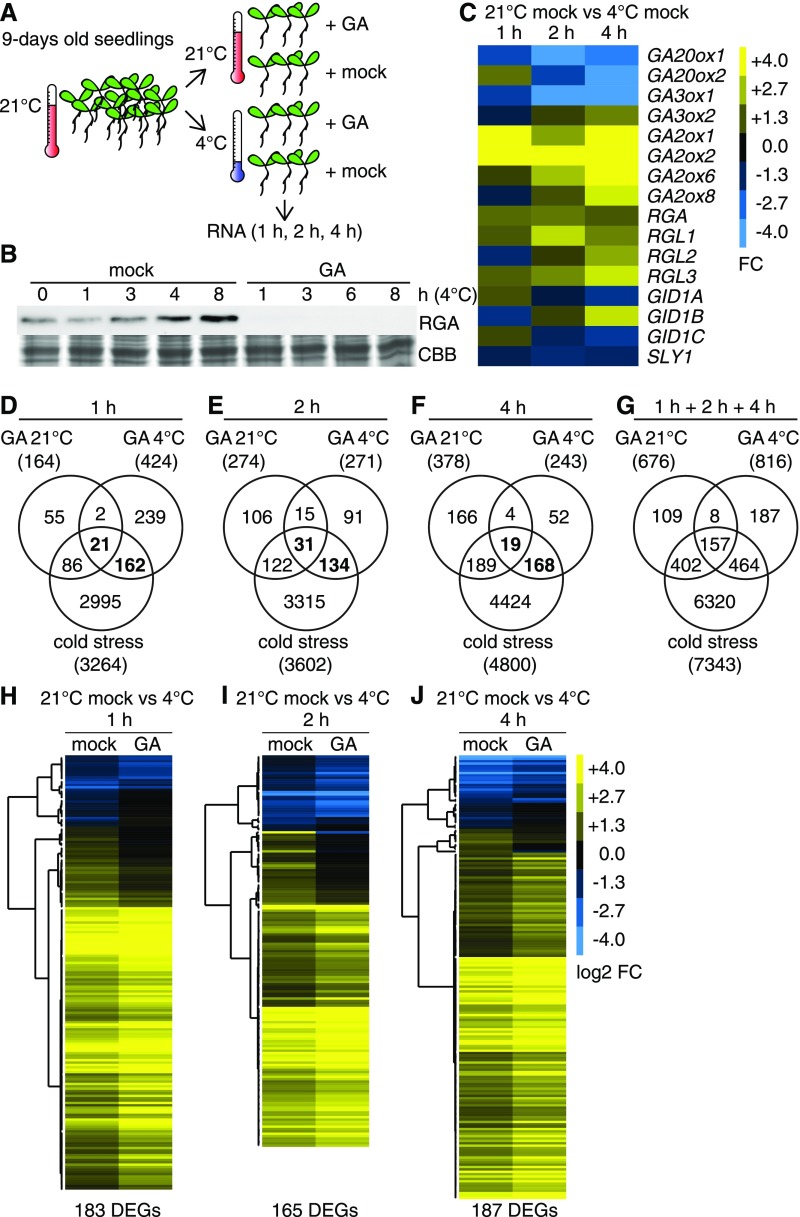
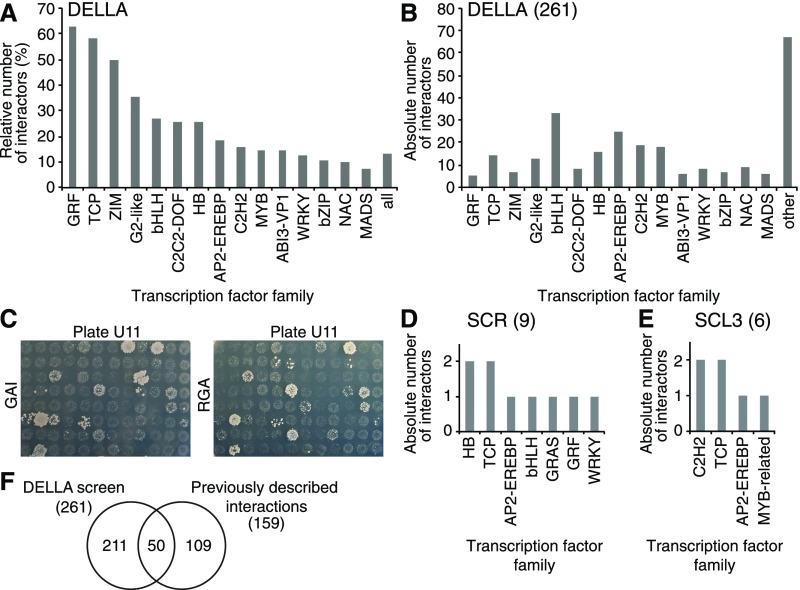
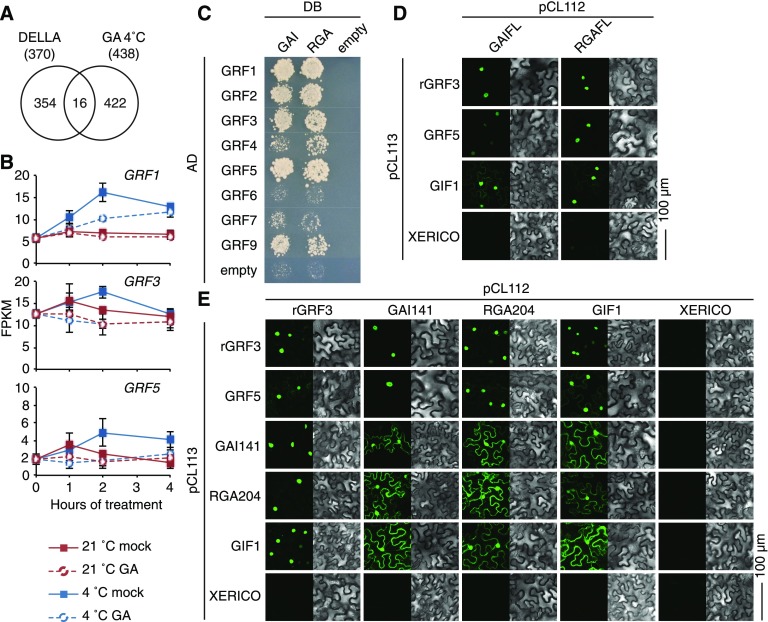
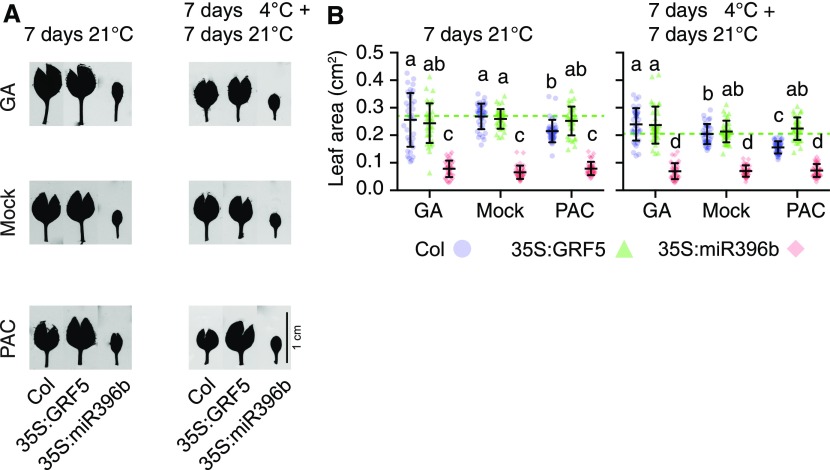
DELLA proteins are repressors of the gibberellin (GA) hormone signaling pathway that act mainly by regulating transcription factor activities in plants. GAs induce DELLA repressor protein degradation and thereby control a number of critical developmental processes as well as responses to stresses such as cold. The strong effect of cold temperatures on many physiological processes has rendered it difficult to assess, based on phenotypic criteria, the role of GA and DELLAs in plant growth during cold stress. Here, we uncover substantial differences in the GA transcriptomes between plants grown at ambient temperature (21°C) and plants exposed to cold stress (4°C) in Arabidopsis (Arabidopsis thaliana). We further identify over 250, to the largest extent previously unknown, DELLA-transcription factor interactions using the yeast two-hybrid system. By integrating both data sets, we reveal that most members of the nine-member GRF (GROWTH REGULATORY FACTOR) transcription factor family are DELLA interactors and, at the same time, that several GRF genes are targets of DELLA-modulated transcription after exposure to cold stress. We find that plants with altered GRF dosage are differentially sensitive to the manipulation of GA and hence DELLA levels, also after cold stress, and identify a subset of cold stress-responsive genes that qualify as targets of this DELLA-GRF regulatory module.
© 2020 American Society of Plant Biologists. All rights reserved.
Figures
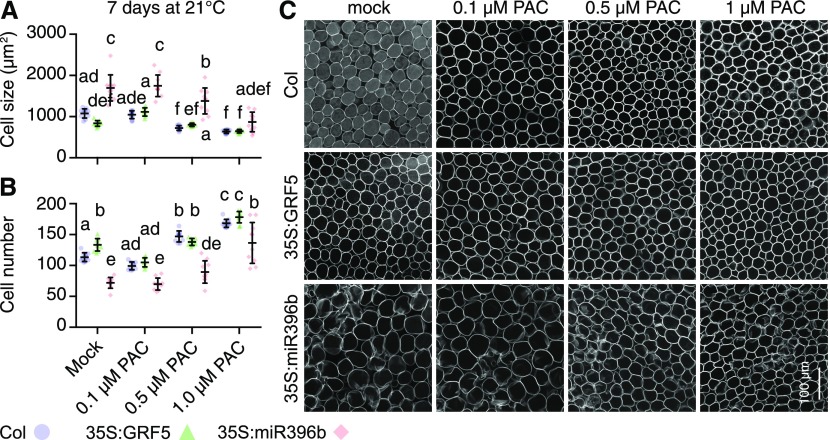
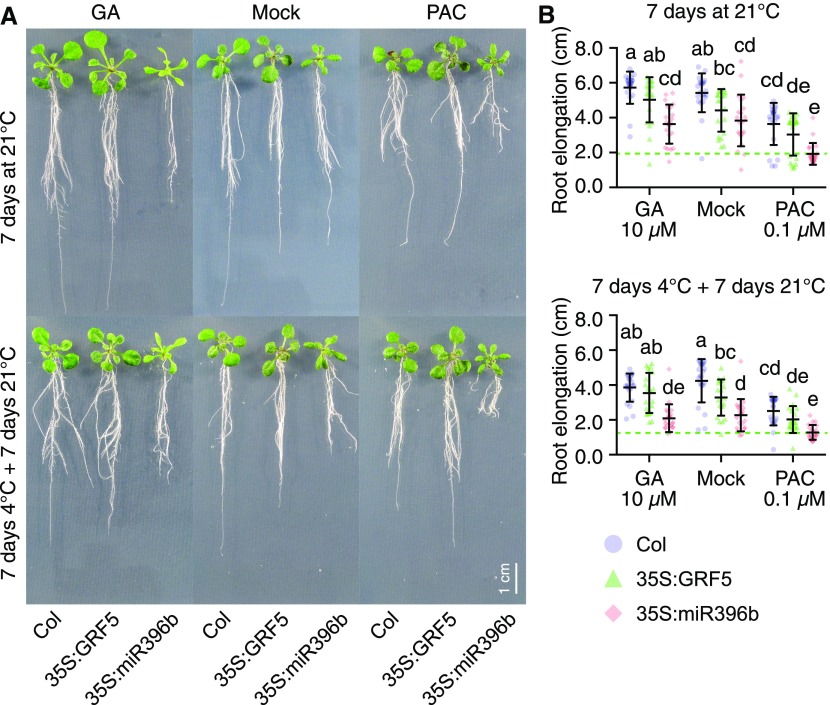
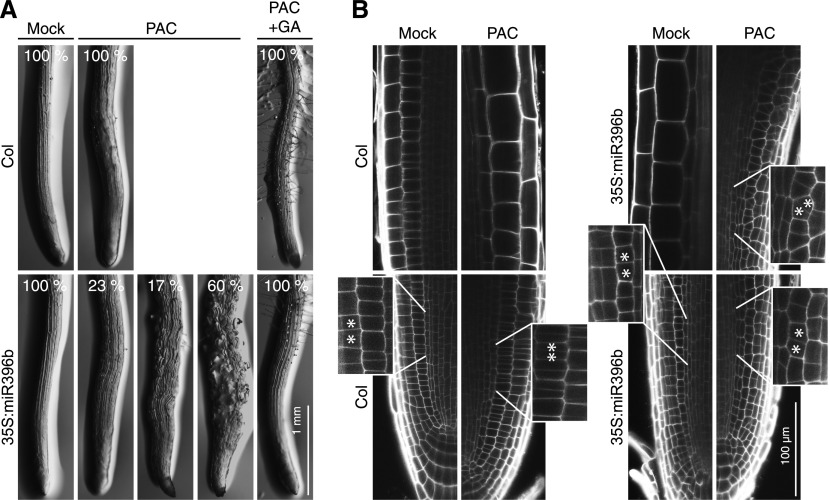
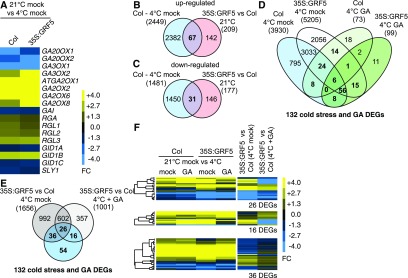
Comment in
-
The Cold Never Bothered Me Anyway: DELLA-Interacting GROWTH REGULATING FACTORS Mediate Plant Growth in Cold Stress.Plant Cell. 2020 Apr;32(4):797-798. doi: 10.1105/tpc.20.00079. Epub 2020 Feb 14. Plant Cell. 2020. PMID: 32060177 Free PMC article. No abstract available.
References
-
- Achard P., Renou J.P., Berthomé R., Harberd N.P., Genschik P.(2008b). Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Curr. Biol. 18: 656–660. - PubMed
-
- Altmann M., Altmann S., Falter C., Falter-Braun P.(2018). High-quality yeast-2-hybrid interaction network mapping. Curr. Protoc. Plant Biol. 3: e20067. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
