

Tracking the homeostasis of second messenger cyclic-di-GMP in bacteria
- PMID: 32060735
- PMCID: PMC7311556
- DOI: 10.1007/s12551-020-00636-1
Tracking the homeostasis of second messenger cyclic-di-GMP in bacteria
Abstract
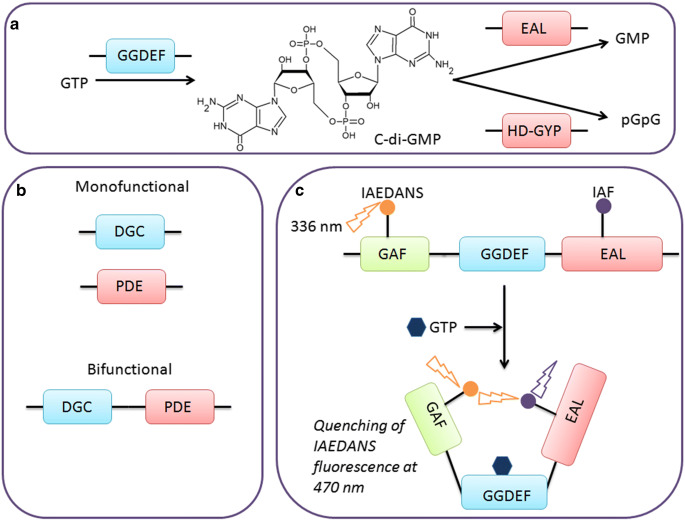
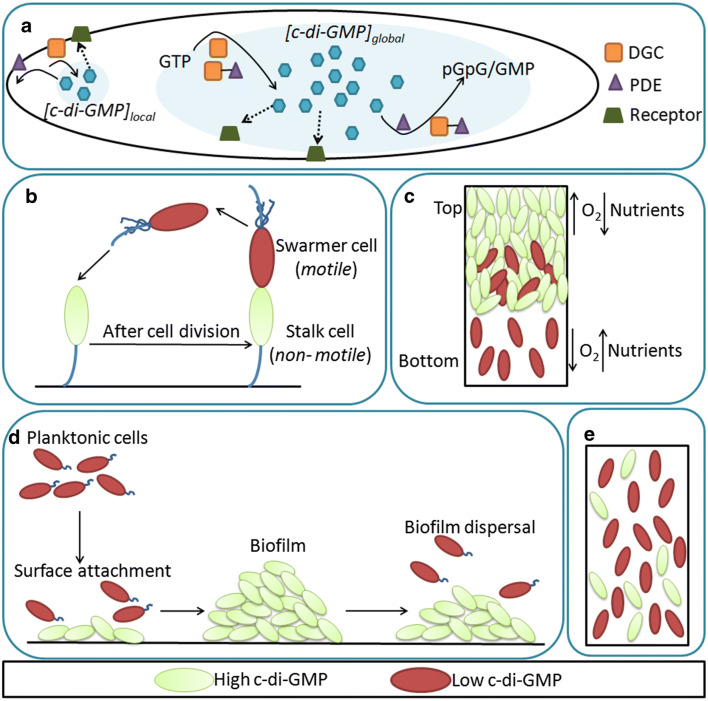
Cyclic-di-GMP (c-di-GMP) is an important second messenger in bacteria which regulates the bacterial transition from motile to sessile phase and also plays a major role in processes such as cell division, exopolysaccharide synthesis, and biofilm formation. Due to its crucial role in dictating the bacterial phenotype, the synthesis and hydrolysis of c-di-GMP is tightly regulated via multiple mechanisms. Perturbing the c-di-GMP homeostasis affects bacterial growth and survival, so it is necessary to understand the underlying mechanisms related to c-di-GMP metabolism. Most techniques used for estimating the c-di-GMP concentration lack single-cell resolution and do not provide information about any heterogeneous distribution of c-di-GMP inside cells. In this review, we briefly discuss how the activity of c-di-GMP metabolising enzymes, particularly bifunctional proteins, is modulated to maintain c-di-GMP homeostasis. We further highlight how fluorescence-based methods aid in understanding the spatiotemporal regulation of c-di-GMP signalling. Finally, we discuss the blind spots in our understanding of second messenger signalling and outline how they can be addressed in the future.
Keywords: Biofilm; Biosensor; C-di-GMP; FRET; Riboswitch; Second messenger.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Amikam D, Weinhouse H, Galperin MY (2010) Moshe Benziman and the discovery of cyclic di-GMP the second messenger cyclic Di-GMP
-
- Bähre H, Kaever V (2017) Identification and quantification of cyclic di-guanosine monophosphate and its linear metabolites by reversed-phase LC-MS/MS. In: c-di-GMP signaling. Springer, pp 45-58 - PubMed
-
- Bharati BK, Sharma IM, Kasetty S, Kumar M, Mukherjee R, Chatterji D (2012) A full-length bifunctional protein involved in c-di-GMP turnover is required for long-term survival under nutrient starvation in Mycobacterium smegmatis Microbiology 158:1415–1427 doi:10.1099/mic.0.053892-0 - PubMed
Publication types
LinkOut - more resources
Full Text Sources
