

Circadian Rhythms in the Pathogenesis and Treatment of Fatty Liver Disease
- PMID: 32061597
- PMCID: PMC7279714
- DOI: 10.1053/j.gastro.2020.01.050
Circadian Rhythms in the Pathogenesis and Treatment of Fatty Liver Disease
Abstract
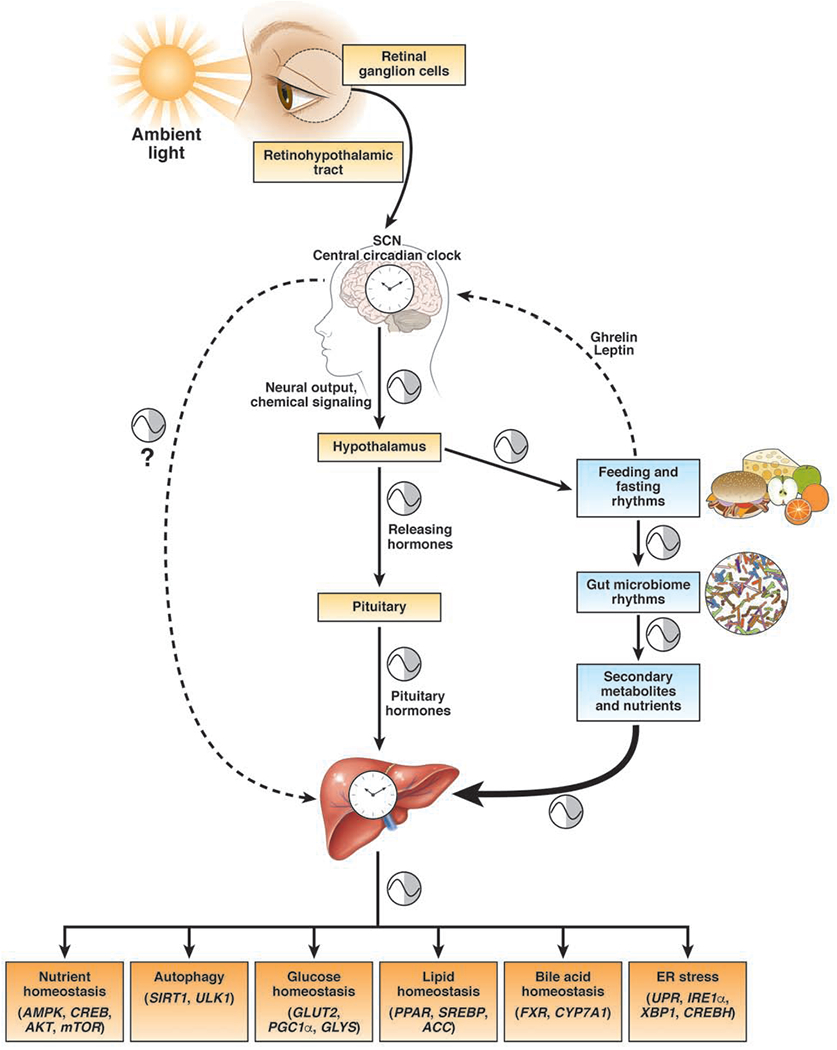
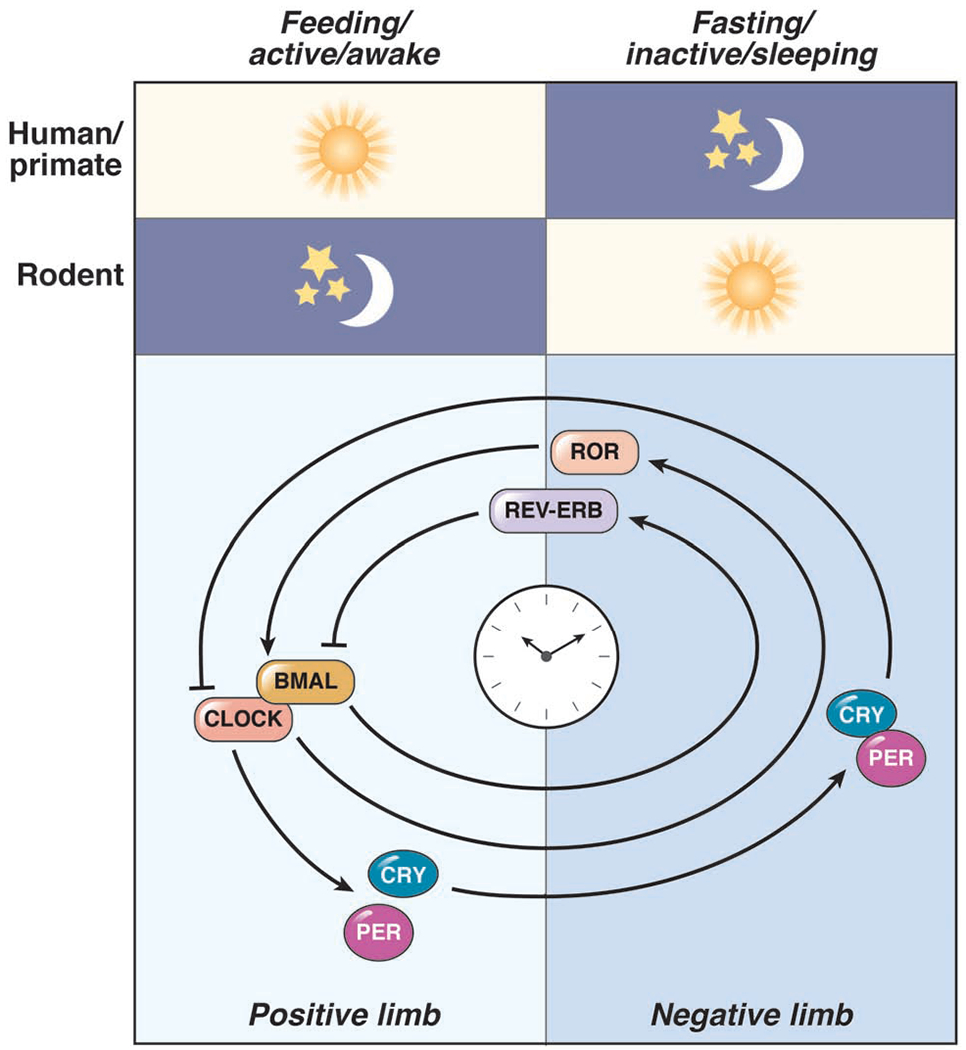
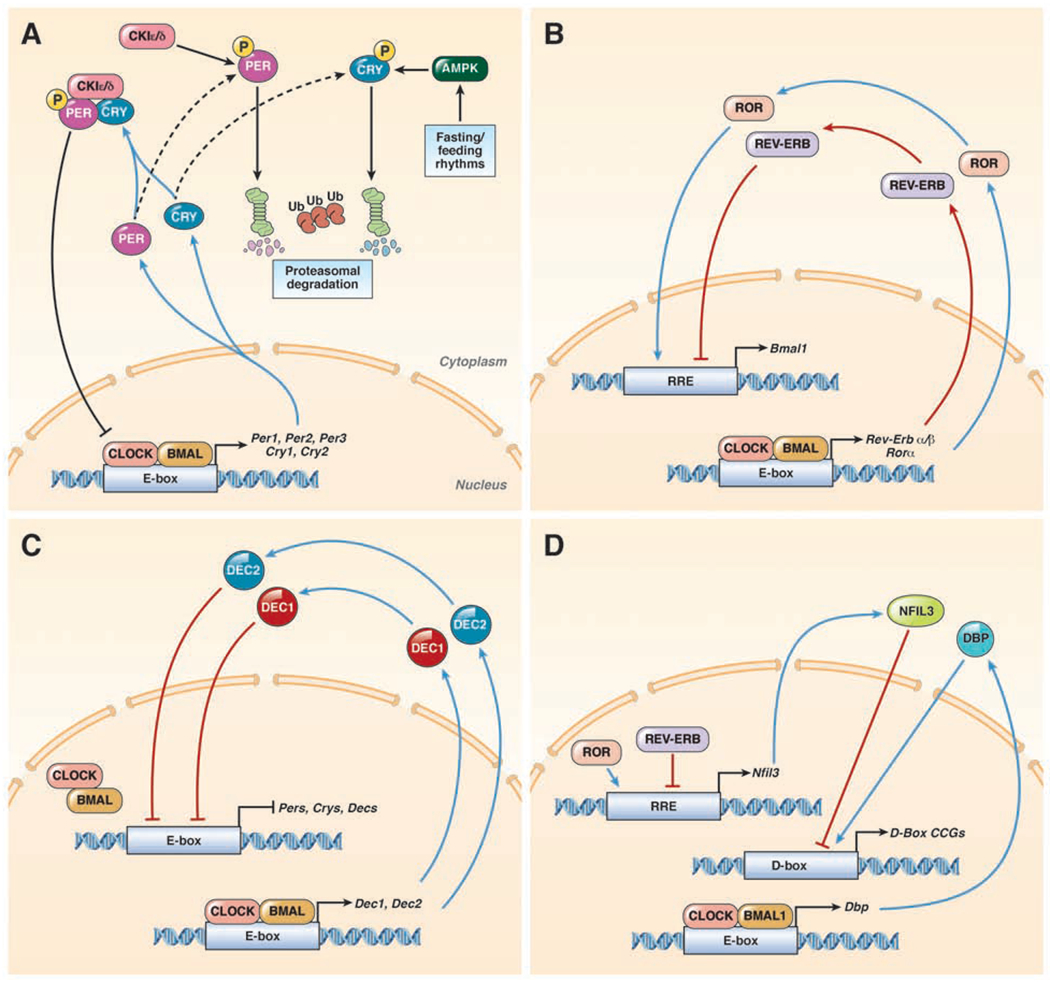
Circadian clock proteins are endogenous timing mechanisms that control the transcription of hundreds of genes. Their integral role in coordinating metabolism has led to their scrutiny in a number of diseases, including nonalcoholic fatty liver disease (NAFLD). Discoordination between central and peripheral circadian rhythms is a core feature of nearly every genetic, dietary, or environmental model of metabolic syndrome and NAFLD. Restricting feeding to a defined daily interval (time-restricted feeding) can synchronize the central and peripheral circadian rhythms, which in turn can prevent or even treat the metabolic syndrome and hepatic steatosis. Importantly, a number of proteins currently under study as drug targets in NAFLD (sterol regulatory element-binding protein [SREBP], acetyl-CoA carboxylase [ACC], peroxisome proliferator-activator receptors [PPARs], and incretins) are modulated by circadian proteins. Thus, the clock can be used to maximize the benefits and minimize the adverse effects of pharmaceutical agents for NAFLD. The circadian clock itself has the potential for use as a target for the treatment of NAFLD.
Keywords: Bile Acids; Dyssynchrony; Gut Microbiome; Lipogenesis; Steatohepatitis.
Published by Elsevier Inc.
Figures
References
-
- Gachon F, Nagoshi E, Brown SA, et al. The mammalian circadian timing system: from gene expression to physiology. Chromosoma 2004;113:103–12. - PubMed
-
- Yamazaki S, Numano R, Abe M, et al. Resetting central and peripheral circadian oscillators in transgenic rats. Science 2000;288:682–5. - PubMed
-
- Buijs R, Salgado R, Sabath E, et al. Peripheral circadian oscillators: time and food. Prog Mol Biol Transl Sci 2013;119:83–103. - PubMed
