

Transcriptome analysis of rat dorsal hippocampal CA1 after an early life seizure induced by kainic acid
- PMID: 32062370
- PMCID: PMC7096153
- DOI: 10.1016/j.eplepsyres.2020.106283
Transcriptome analysis of rat dorsal hippocampal CA1 after an early life seizure induced by kainic acid
Abstract
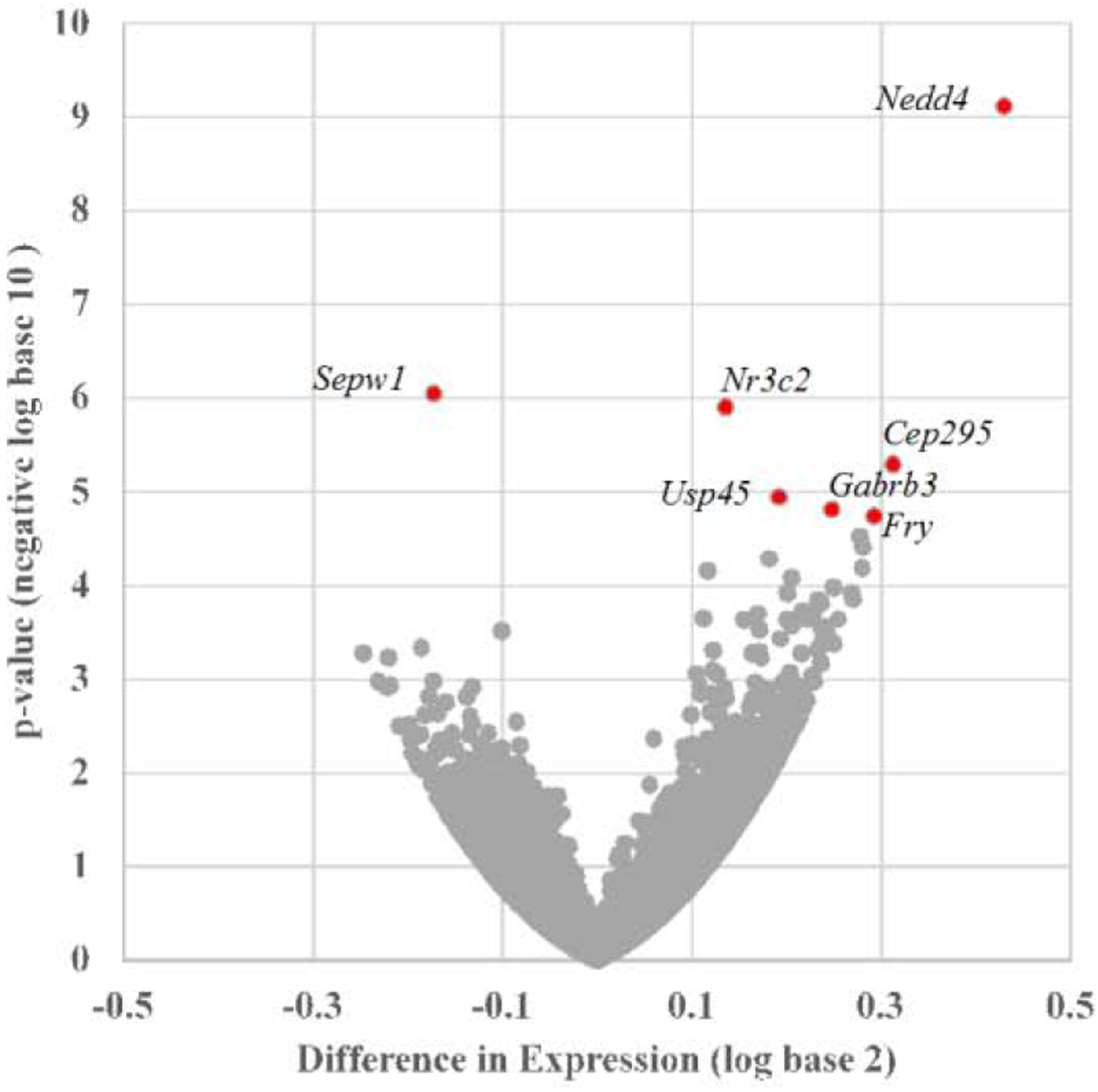
Seizures that occur during early development are associated with adverse neurodevelopmental outcomes. Causation and mechanisms are currently under investigation. Induction of an early life seizure by kainic acid (KA) in immature rats on post-natal day (P) 7 results in behavioral changes in the adult rat that reflect social and intellectual deficits without overt cellular damage. Our previous work also demonstrated increased expression of CA1 hippocampal long-term potentiation (LTP) and reduced desensitization of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid-type ionotropic glutamate receptors (AMPA-R) one week following a kainic acid induced seizure (KA-ELS). Here we used RNA sequencing (RNAseq) of mRNA from dorsal hippocampal CA1 to probe changes in mRNA levels one week following KA-ELS as a means to investigate the mechanisms for these functional changes. Ingenuity pathway analysis (IPA) confirmed our previous results by predicting an up-regulation of the synaptic LTP pathway. Differential gene expression results revealed significant differences in 7 gene isoforms. Additional assessments included AMPA-R splice variants and adenosine deaminase acting on RNA 2 (ADAR2) editing sites as a means to determine the mechanism for reduced AMPA-R desensitization. Splice variant analysis demonstrated that KA-ELS result in a small, but significant decrease in the "flop" isoform of Gria3, and editing site analysis revealed significant changes in the editing of a kainate receptor subunit, Grik2, and a serotonin receptor, Htr2c. While these specific changes may not account for altered AMPA-R desensitization, the differences indicate that KA-ELS alters gene expression in the hippocampal CA1 one week after the insult.
Keywords: ADAR2; Development; Kainate; RNAseq; Seizure; Splice-variant.
Copyright © 2020 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare no financial or non-financial competing interests.
Figures



References
-
- Aken BL, Achuthan P, Akanni W, Amode MR, Bernsdorff F, Bhai J, Billis K, Carvalho-Silva D, Cummins C, Clapham P, Gil L, Giron CG, Gordon L, Hourlier T, Hunt SE, Janacek SH, Juettemann T, Keenan S, Laird MR, Lavidas I, Maurel T, McLaren W, Moore B, Murphy DN, Nag R, Newman V, Nuhn M, Ong CK, Parker A, Patricio M, Riat HS, Sheppard D, Sparrow H, Taylor K, Thormann A, Vullo A, Walts B, Wilder SP, Zadissa A, Kostadima M, Martin FJ, Muffato M, Perry E, Ruffier M, Staines DM, Trevanion SJ, Cunningham F, Yates A, Zerbino DR, Flicek P, 2017. Ensembl 2017. Nucleic Acids Res 45, D635–D642. - PMC - PubMed
-
- Amantana A, Vorachek WR, Butler JA, Costa ND, Whanger PD, 2002. Effect of copper, zinc and cadmium on the promoter of selenoprotein W in glial and myoblast cells. J Inorg Biochem 91, 356–362. - PubMed
-
- Bakker CE, Oostra BA, 2003. Understanding fragile X syndrome: insights from animal models. Cytogenet Genome Res 100, 111–123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
