

Why leveraging sex differences in immune trade-offs may illuminate the evolution of senescence
- PMID: 32063662
- PMCID: PMC7006808
- DOI: 10.1111/1365-2435.13458
Why leveraging sex differences in immune trade-offs may illuminate the evolution of senescence
Abstract
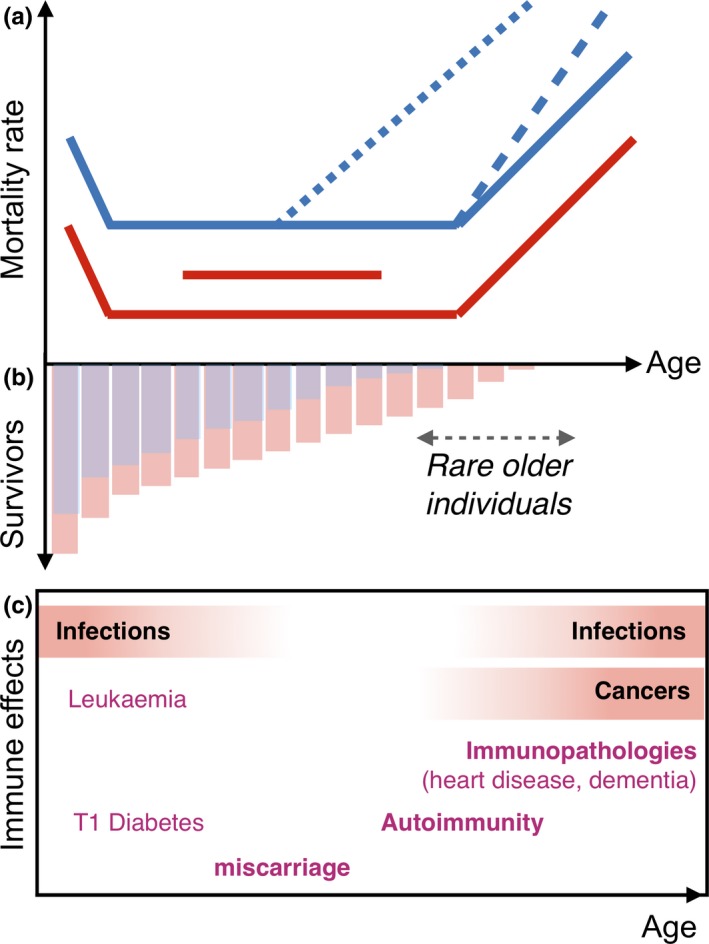
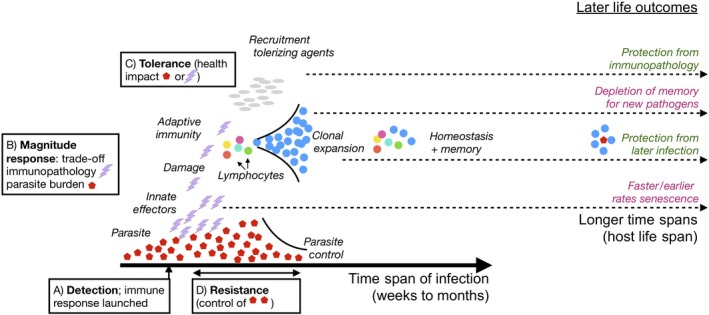
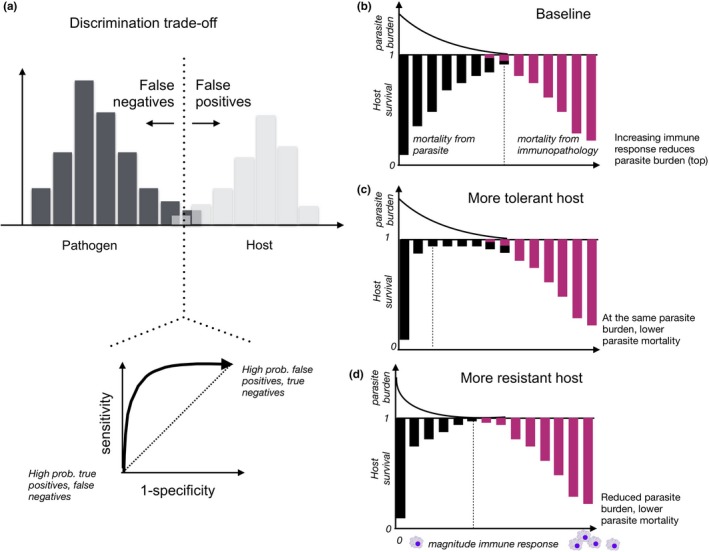
The immune system affects senescence (declines in probabilities of survival or reproduction with age), by shaping late age vulnerability to chronic inflammatory diseases and infections. It is also a dynamic interactive system that must balance competing demands across the life course. Thus, immune system function remains an important frontier in understanding the evolution of senescence.Here, we review our expanding mechanistic understanding of immune function over the life course, in the context of theoretical predictions from life-history evolution. We are especially interested in stage- and sex-dependent costs and benefits of investment in the immune system, given differential life-history priorities of the life stages and sexes.We introduce the costs likely to govern immune allocation across the life course. We then discuss theoretical expectations for differences between the sexes and their likely consequences in terms of how the immune system is both modulated by and may modulate senescence, building on information from life-history theory, experimental immunology and demography.We argue that sex differences in immune function provide a potentially powerful probe of selection pressures on the immune system across the life course. In particular, differences in 'competing' and 'caring' between the sexes have evolved across the tree of life, providing repeated instances of divergent selection pressures on immune function occurring within the same overall bauplan.We conclude by detailing an agenda for future research, including development of theoretical predictions of the differences between the sexes under an array of existing models for sex differences in immunity, and empirical tests of such predictions across the tree of life. A free http://onlinelibrary.wiley.com/doi/10.1111/1365-2435.13458/suppinfo can be found within the Supporting Information of this article.
Keywords: immunity; life history theory; senescence; sex differences.
© 2019 The Authors. Functional Ecology published by John Wiley & Sons Ltd on behalf of British Ecological Society.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources