

miRNA-31 Improves Cognition and Abolishes Amyloid-β Pathology by Targeting APP and BACE1 in an Animal Model of Alzheimer's Disease
- PMID: 32069773
- PMCID: PMC7031139
- DOI: 10.1016/j.omtn.2020.01.010
miRNA-31 Improves Cognition and Abolishes Amyloid-β Pathology by Targeting APP and BACE1 in an Animal Model of Alzheimer's Disease
Abstract
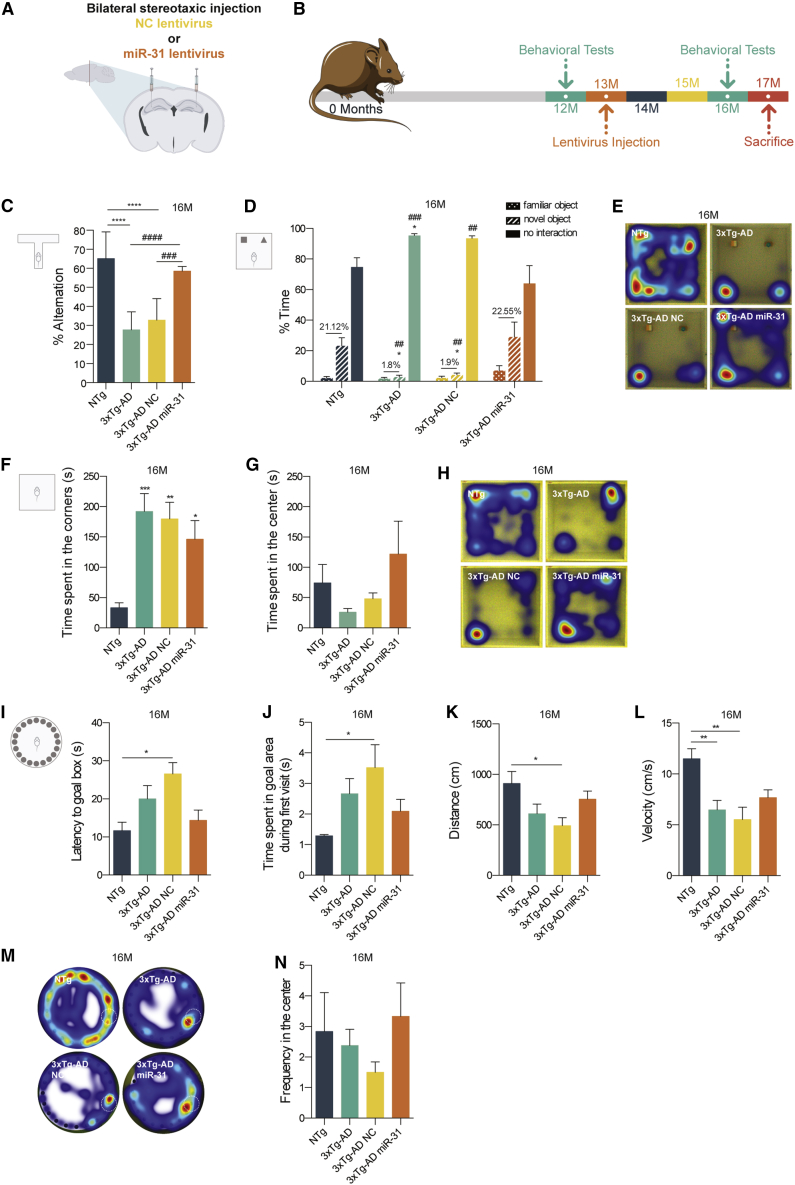
Alzheimer's disease (AD) is the most common form of dementia worldwide, characterized by progressive memory impairment, behavioral changes, and, ultimately, loss of consciousness and death. Recently, microRNA (miRNA) dysfunction has been associated with increased production and impaired clearance of amyloid-β (Aβ) peptides, whose accumulation is one of the most well-known pathophysiological markers of this disease. In this study, we identified several miRNAs capable of targeting key proteins of the amyloidogenic pathway. The expression of one of these miRNAs, miR-31, previously found to be decreased in AD patients, was able to simultaneously reduce the levels of APP and Bace1 mRNA in the hippocampus of 17-month-old AD triple-transgenic (3xTg-AD) female mice, leading to a significant improvement of memory deficits and a reduction in anxiety and cognitive inflexibility. In addition, lentiviral-mediated miR-31 expression significantly ameliorated AD neuropathology in this model, drastically reducing Aβ deposition in both the hippocampus and subiculum. Furthermore, the increase of miR-31 levels was enough to reduce the accumulation of glutamate vesicles in the hippocampus to levels found in non-transgenic age-matched animals. Overall, our results suggest that miR-31-mediated modulation of APP and BACE1 can become a therapeutic option in the treatment of AD.
Keywords: APP; Alzheimer’s disease; BACE1; amyloid-β peptide; cognitive function; gene therapy; lentiviral vector; memory; miR-31.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures






References
-
- Goldman J.S., Hahn S.E., Catania J.W., LaRusse-Eckert S., Butson M.B., Rumbaugh M., Strecker M.N., Roberts J.S., Burke W., Mayeux R., Bird T., American College of Medical Genetics and the National Society of Genetic Counselors Genetic counseling and testing for Alzheimer disease: joint practice guidelines of the American College of Medical Genetics and the National Society of Genetic Counselors. Genet. Med. 2011;13:597–605. - PMC - PubMed
-
- Hardy J.A., Higgins G.A. Alzheimer’s disease: the amyloid cascade hypothesis. Science. 1992;256:184–185. - PubMed
-
- Karran E., Mercken M., De Strooper B. The amyloid cascade hypothesis for Alzheimer’s disease: an appraisal for the development of therapeutics. Nat. Rev. Drug Discov. 2011;10:698–712. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
