

Prospective isolation of chondroprogenitors from human iPSCs based on cell surface markers identified using a CRISPR-Cas9-generated reporter
- PMID: 32070421
- PMCID: PMC7026983
- DOI: 10.1186/s13287-020-01597-8
Prospective isolation of chondroprogenitors from human iPSCs based on cell surface markers identified using a CRISPR-Cas9-generated reporter
Abstract
Background: Articular cartilage shows little or no capacity for intrinsic repair, generating a critical need of regenerative therapies for joint injuries and diseases such as osteoarthritis. Human-induced pluripotent stem cells (hiPSCs) offer a promising cell source for cartilage tissue engineering and in vitro human disease modeling; however, off-target differentiation remains a challenge during hiPSC chondrogenesis. Therefore, the objective of this study was to identify cell surface markers that define the true chondroprogenitor population and use these markers to purify iPSCs as a means of improving the homogeneity and efficiency of hiPSC chondrogenic differentiation.
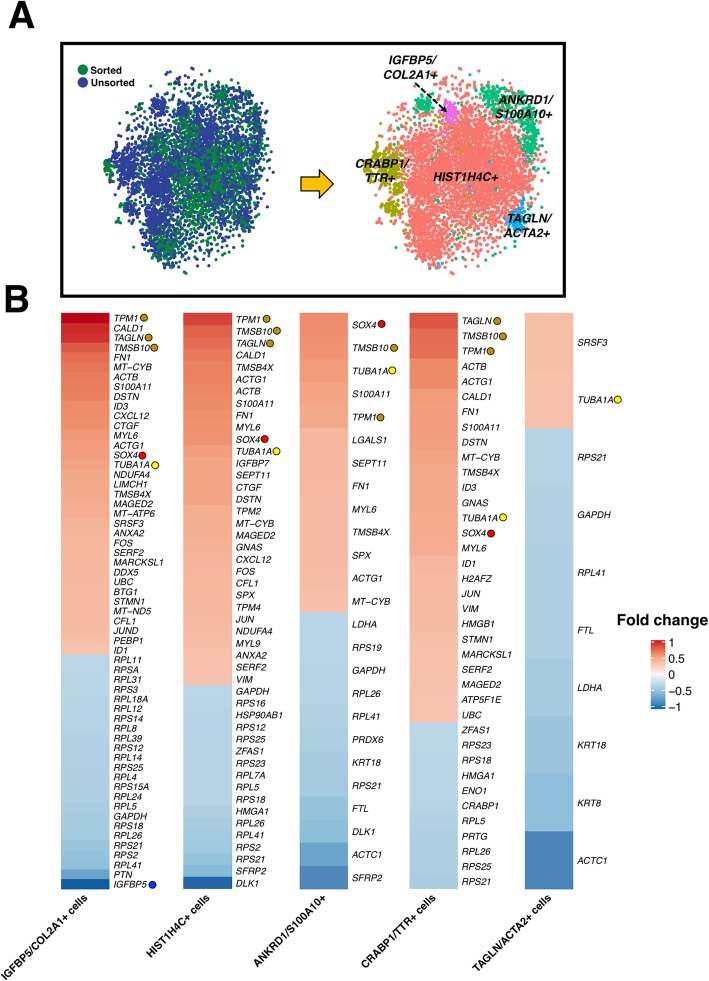
Methods: We used a CRISPR-Cas9-edited COL2A1-GFP knock-in reporter hiPSC line, coupled with a surface marker screen, to identify a novel chondroprogenitor population. Single-cell RNA sequencing was then used to analyze the distinct clusters within the population. An unpaired t test with Welch's correction or an unpaired Kolmogorov-Smirnov test was performed with significance reported at a 95% confidence interval.
Results: Chondroprogenitors expressing CD146, CD166, and PDGFRβ, but not CD45, made up an average of 16.8% of the total population. Under chondrogenic culture conditions, these triple-positive chondroprogenitor cells demonstrated decreased heterogeneity as measured by single-cell RNA sequencing with fewer clusters (9 clusters in unsorted vs. 6 in sorted populations) closer together. Additionally, there was more robust and homogenous matrix production (unsorted: 1.5 ng/ng vs. sorted: 19.9 ng/ng sGAG/DNA; p < 0.001) with significantly higher chondrogenic gene expression (i.e., SOX9, COL2A1, ACAN; p < 0.05).
Conclusions: Overall, this study has identified a unique hiPSC-derived subpopulation of chondroprogenitors that are CD146+/CD166+/PDGFRβ+/CD45- and exhibit high chondrogenic potential, providing a purified cell source for cartilage tissue engineering or disease modeling studies.
Keywords: Cartilage; Chondrocyte; Chondroprogenitor; Differentiation; Single-cell RNA sequencing; Surface markers; hiPSC.
Conflict of interest statement
Dr. Guilak is an employee of Cytex Therapeutics, Inc. The other authors declare that they have no competing interests.
Figures





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
