

Review
doi: 10.1104/pp.19.01310.
Epub 2020 Feb 18.
Twenty Years of Progress in Physiological and Biochemical Investigation of RALF Peptides
Affiliations
- PMID: 32071151
- PMCID: PMC7140910
- DOI: 10.1104/pp.19.01310
Item in Clipboard
Review
Twenty Years of Progress in Physiological and Biochemical Investigation of RALF Peptides
Plant Physiol.
2020 Apr.
Abstract
RALF isoforms play many biological roles, and their specific functions are defined by combinatorial interactions with dynamic receptor complexes that vary more than initially thought.
Figures
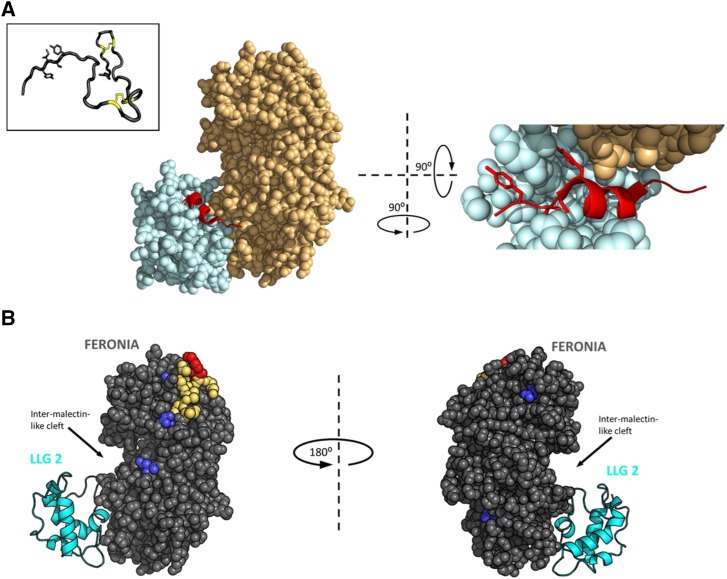
RALF peptides adopt an induced fold when complexed with a CrRLK1L protein. A, the left shows the crystal structure of the heterotypic complex among LLG2 (pale blue), RALF23 (red), and the ectodomain of FERONIA (pale orange). Density for the full-length RALF23 peptide was obtained for residues from RYISY to the first conserved Cys of the mature peptide. The right shows 900 pivot along the y axis by an ∼900 tilt out of the page and zoomed in, with ∼40 residues of the N-terminal domain of ectoFER removed to clear the view of RALF23. The mature peptide hormone intercalates tightly between the juxtamembrane domain of FERONIA (light orange) and the flexible loops of LLG2 (light blue), adopting an induced alpha helix. YISY sidechains are shown as sticks for emphasis. PDB ID: 6A5E (Xiao et al., 2019). The inset shows the solution NMR structure of RALF8 confirms a mostly unstructured mature peptide with a minimal loop-spanning disulfide configuration formed from the four invariant cysteines. Bridged Cys sidechains are represented as yellow sticks. YITY sidechains are shown as sticks for emphasis. PDB ID: 6NU4 (Frederick et al., 2019). B, Chemical cross linking and footprinting data from RALF1 complexed with the extracellular domain of FERONIA (gray spheres) in vitro, in the absence of an LLG protein, mapped to the crystal structure 6A5E. LLG2 (cyan cartoon) is included in the image for spatial reference but was not part of the experiment. Dark blue, residues that gain protection from the bulk solvent in the presence of full-length RALF1 (EDC-GEE labeling); red, Lys 60 (K60) on FERONIA, which cross linked to the C-terminal Lys of RALF1 (BS3 cross linking); yellow, the peptide containing K60, identified by mass spectrometry (Liu et al., 2018).
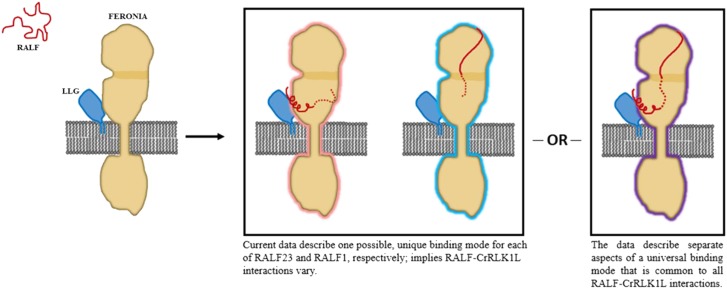
Two proposed models considering available data on the induced fold of AtRALF and AtRALF-CrRLK1L protein binding interactions. To date, the solution NMR structure of AtRALF8 (Frederick et al., 2019) remains the only structure of an unbound AtRALF peptide and suggests that most Cys-rich peptides are similarly disordered. Note that LLG is shown here associated with FER before AtRALF binding for simplicity, but direct experimental evidence of AtRALF-independent LLG-FER interactions has not yet been demonstrated. Model 1 shows ehe crystal structure of the ectodomain of FERONIA complexed with LLG2 and AtRALF23 is currently the only structure of an AtRALF-receptor complex and suggests that the N terminus of AtRALF23 assumes an induced fold during binding and that its YISY motif is highly involved in AtRALF binding (Xiao et al., 2019). Meanwhile, the chemical cross linking and protein footprinting study of the AtRALF1/FER complex in the absence of an LLG protein suggests that the C terminus of AtRALF1 spans the intermalectin-like cleft. In the case of AtRALF23, binding occurs in an LLG-dependent manner near the juxtamembrane domain while AtRALF1 binding occurs independently of LLG at the malectin-like domains, suggesting two distinct, AtRALF-specific binding mechanisms. Model 2 shows, alternatively, these two binding modes reflect an interaction that is generalizable to many mature AtRALF peptides. Until more comprehensive data are available on AtRALF-receptor interactions, this remains a possibility.


References
-
- Belkhadir Y, Jaillais Y(2015) The molecular circuitry of brassinosteroid signaling. New Phytol 206: 522–540 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
