

Target gene-independent functions of MYC oncoproteins
- PMID: 32071436
- PMCID: PMC7611238
- DOI: 10.1038/s41580-020-0215-2
Target gene-independent functions of MYC oncoproteins
Abstract
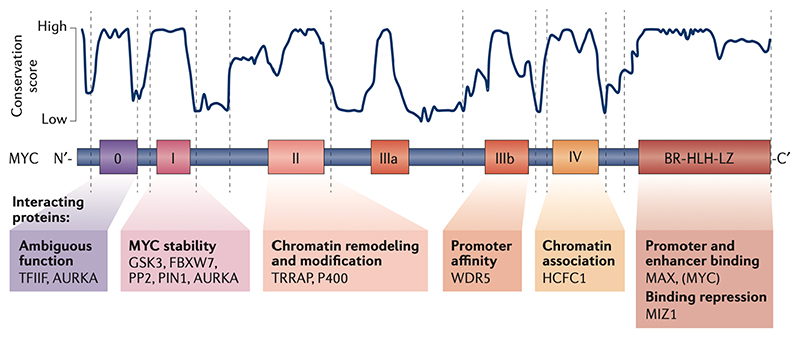
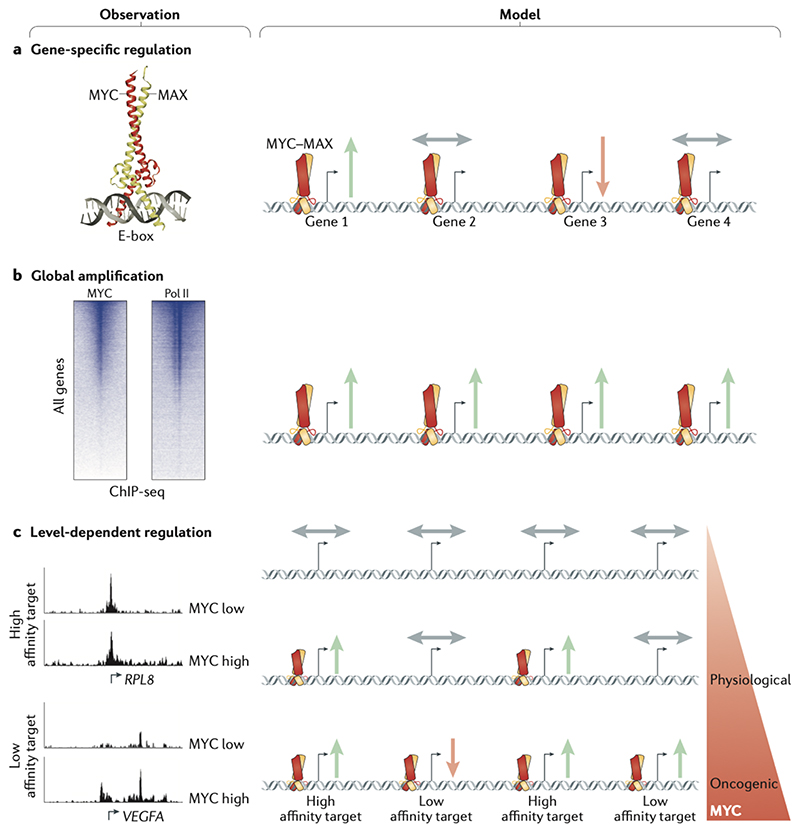
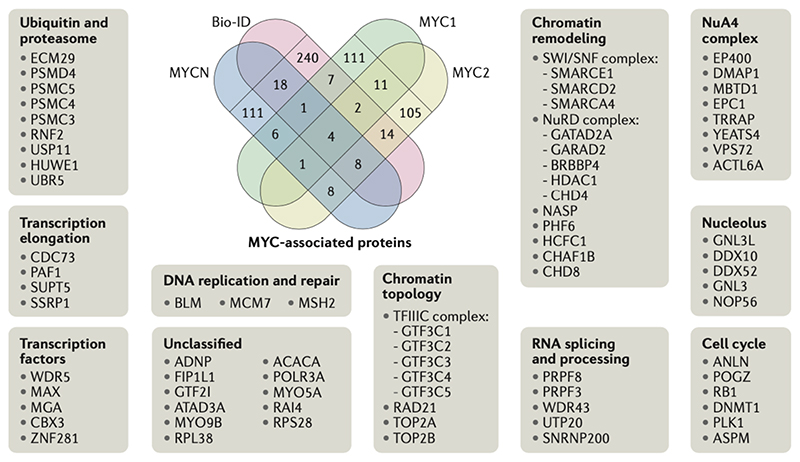
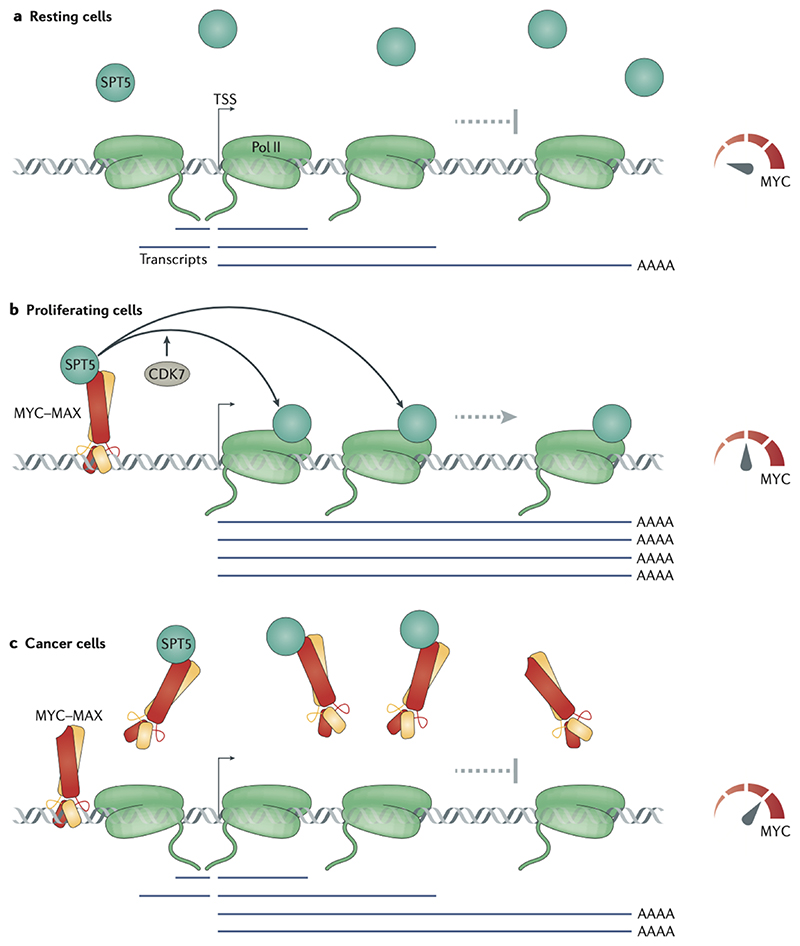
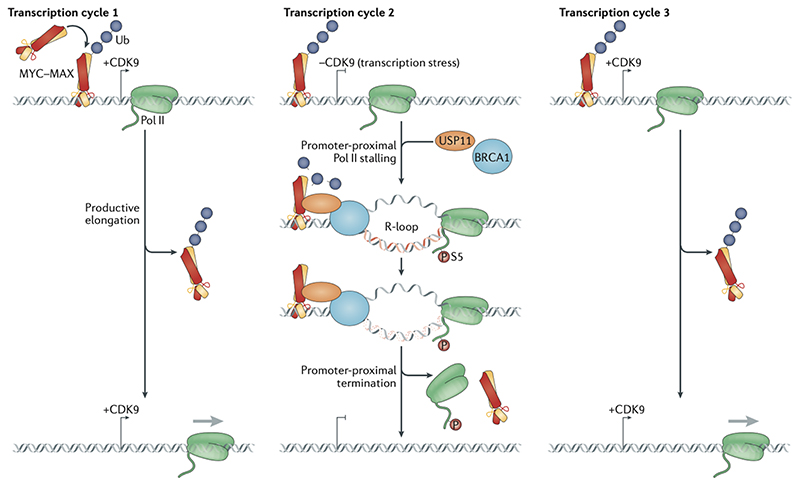
Oncoproteins of the MYC family are major drivers of human tumorigenesis. Since a large body of evidence indicates that MYC proteins are transcription factors, studying their function has focused on the biology of their target genes. Detailed studies of MYC-dependent changes in RNA levels have provided contrasting models of the oncogenic activity of MYC proteins through either enhancing or repressing the expression of specific target genes, or as global amplifiers of transcription. In this Review, we first summarize the biochemistry of MYC proteins and what is known (or is unclear) about the MYC target genes. We then discuss recent progress in defining the interactomes of MYC and MYCN and how this information affects central concepts of MYC biology, focusing on mechanisms by which MYC proteins modulate transcription. MYC proteins promote transcription termination upon stalling of RNA polymerase II, and we propose that this mechanism enhances the stress resilience of basal transcription. Furthermore, MYC proteins coordinate transcription elongation with DNA replication and cell cycle progression. Finally, we argue that the mechanism by which MYC proteins regulate the transcription machinery is likely to promote tumorigenesis independently of global or relative changes in the expression of their target genes.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
