

An atlas of transposable element-derived alternative splicing in cancer
- PMID: 32075558
- PMCID: PMC7061986
- DOI: 10.1098/rstb.2019.0342
An atlas of transposable element-derived alternative splicing in cancer
Abstract
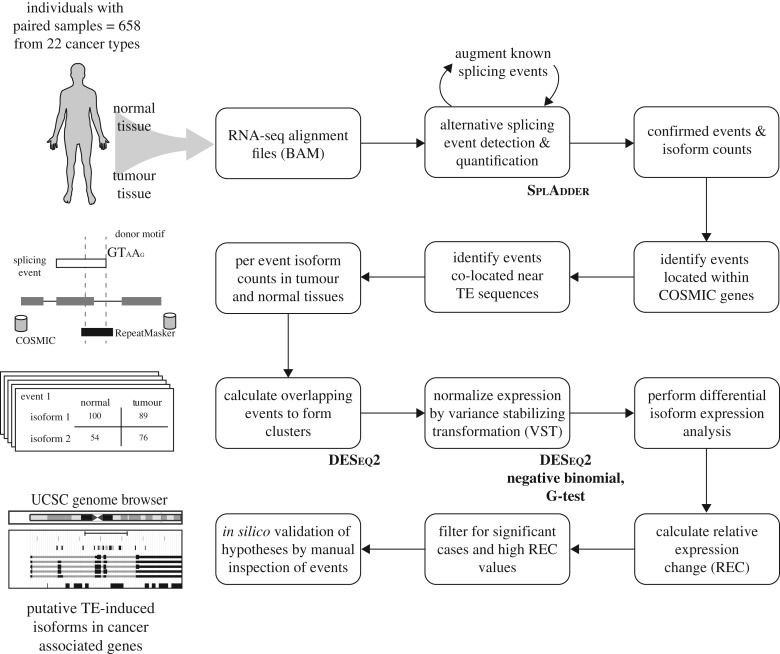
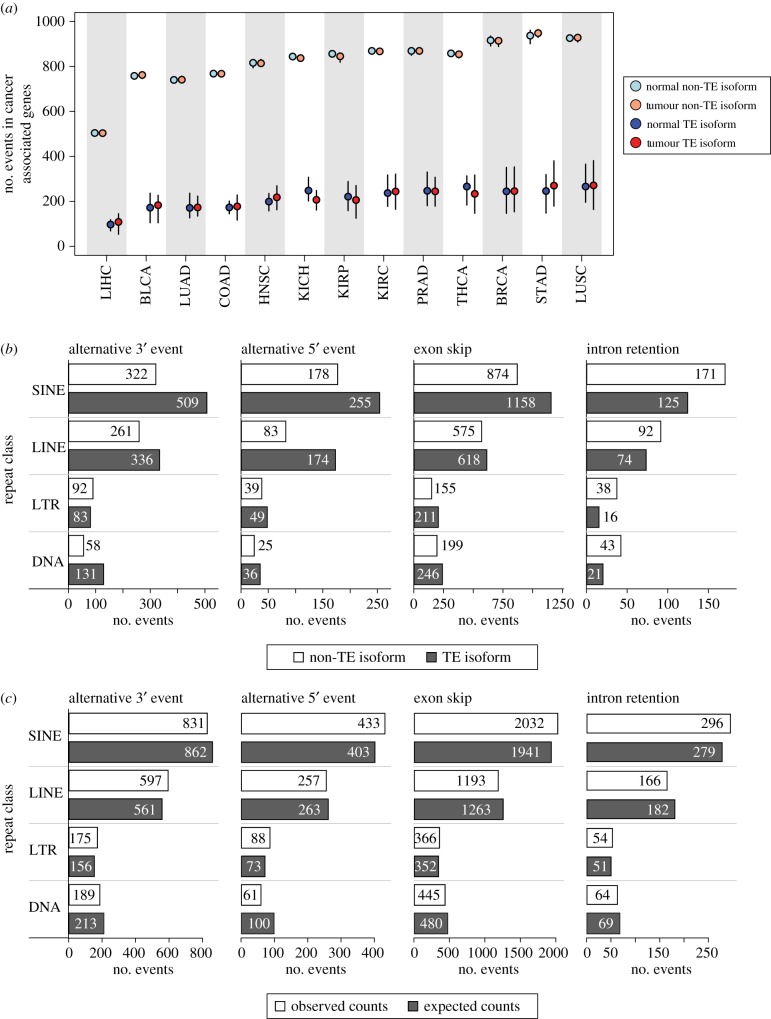
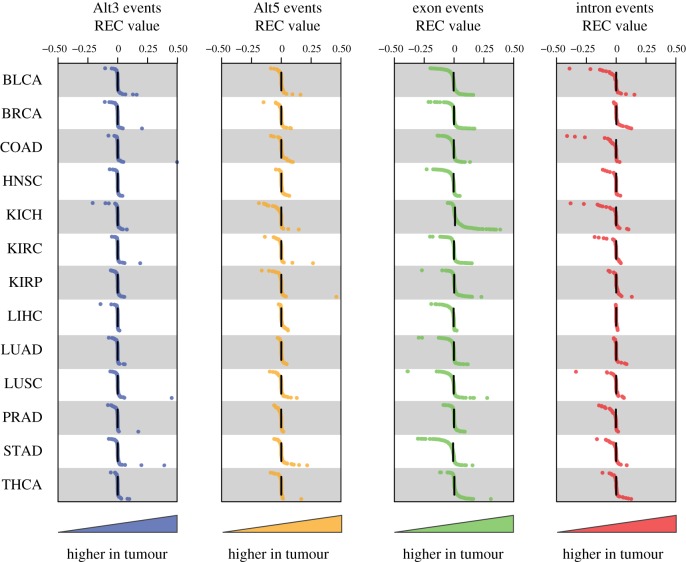
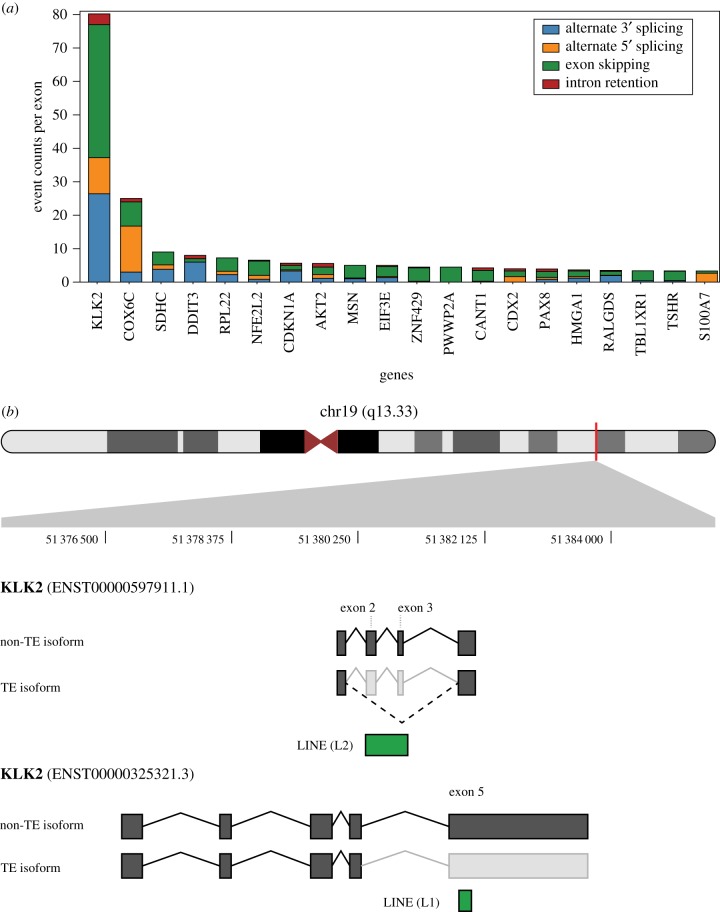
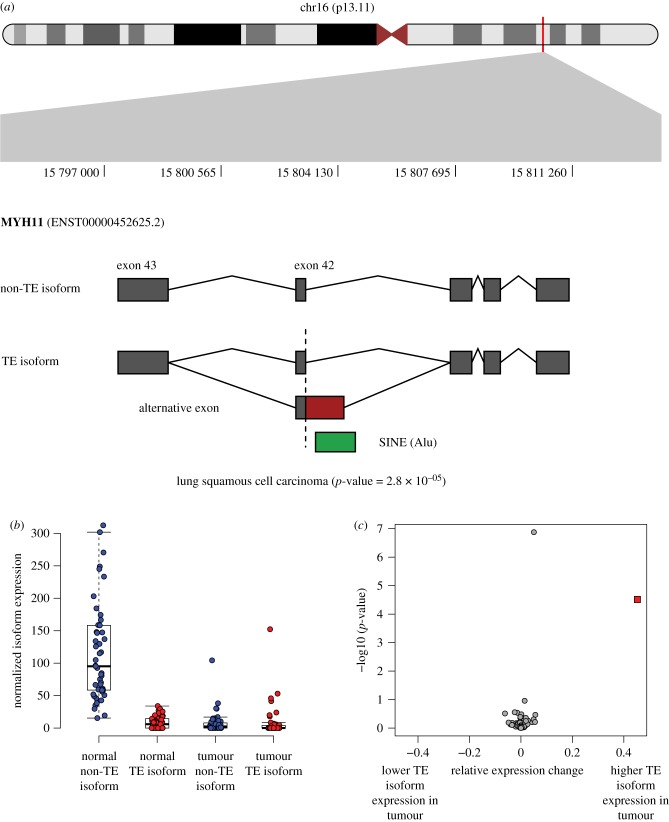
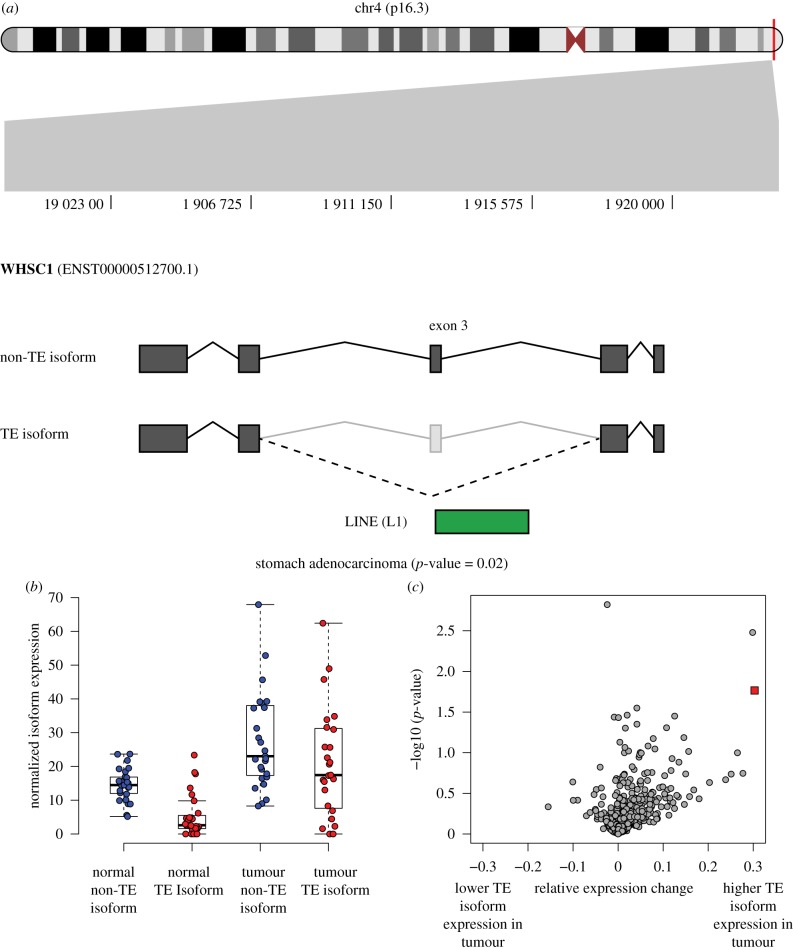
Transposable element (TE)-derived sequences comprise more than half of the human genome, and their presence has been documented to alter gene expression in a number of different ways, including the generation of alternatively spliced transcript isoforms. Alternative splicing has been associated with tumorigenesis for a number of different cancers. The objective of this study was to broadly characterize the role of human TEs in generating alternatively spliced transcript isoforms in cancer. To do so, we screened for the presence of TE-derived sequences co-located with alternative splice sites that are differentially used in normal versus cancer tissues. We analysed a comprehensive set of alternative splice variants characterized for 614 matched normal-tumour tissue pairs across 13 cancer types, resulting in the discovery of 4820 TE-generated alternative splice events distributed among 723 cancer-associated genes. Short interspersed nuclear elements (Alu) and long interspersed nuclear elements (L1) were found to contribute the majority of TE-generated alternative splice sites in cancer genes. A number of cancer-associated genes, including MYH11, WHSC1 and CANT1, were shown to have overexpressed TE-derived isoforms across a range of cancer types. TE-derived isoforms were also linked to cancer-specific fusion transcripts, suggesting a novel mechanism for the generation of transcriptome diversity via trans-splicing mediated by dispersed TE repeats. This article is part of a discussion meeting issue 'Crossroads between transposons and gene regulation'.
Keywords: alternative splicing; cancer; gene expression; gene regulation; transposable elements; tumorigenesis.
Conflict of interest statement
We declare we have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical