

Sen1 Is Recruited to Replication Forks via Ctf4 and Mrc1 and Promotes Genome Stability
- PMID: 32075754
- PMCID: PMC7034062
- DOI: 10.1016/j.celrep.2020.01.087
Sen1 Is Recruited to Replication Forks via Ctf4 and Mrc1 and Promotes Genome Stability
Abstract
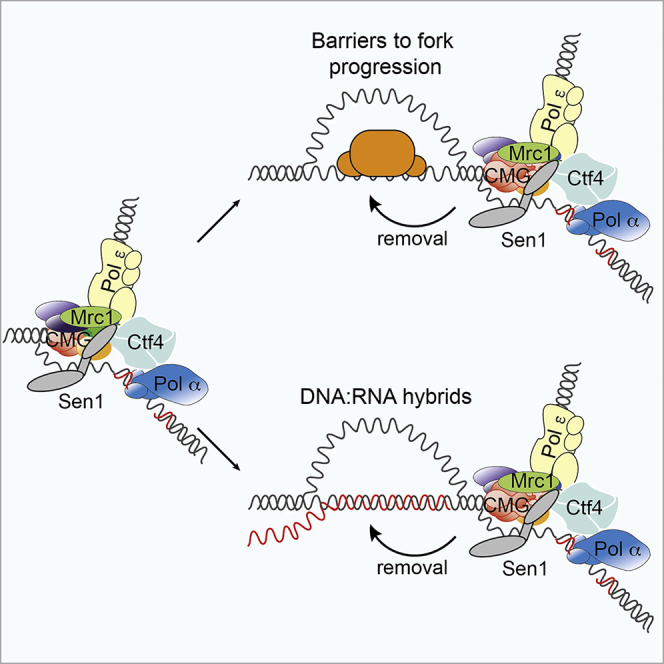
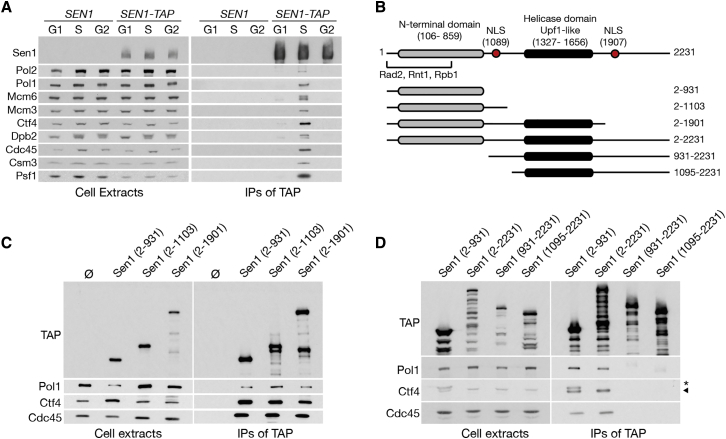
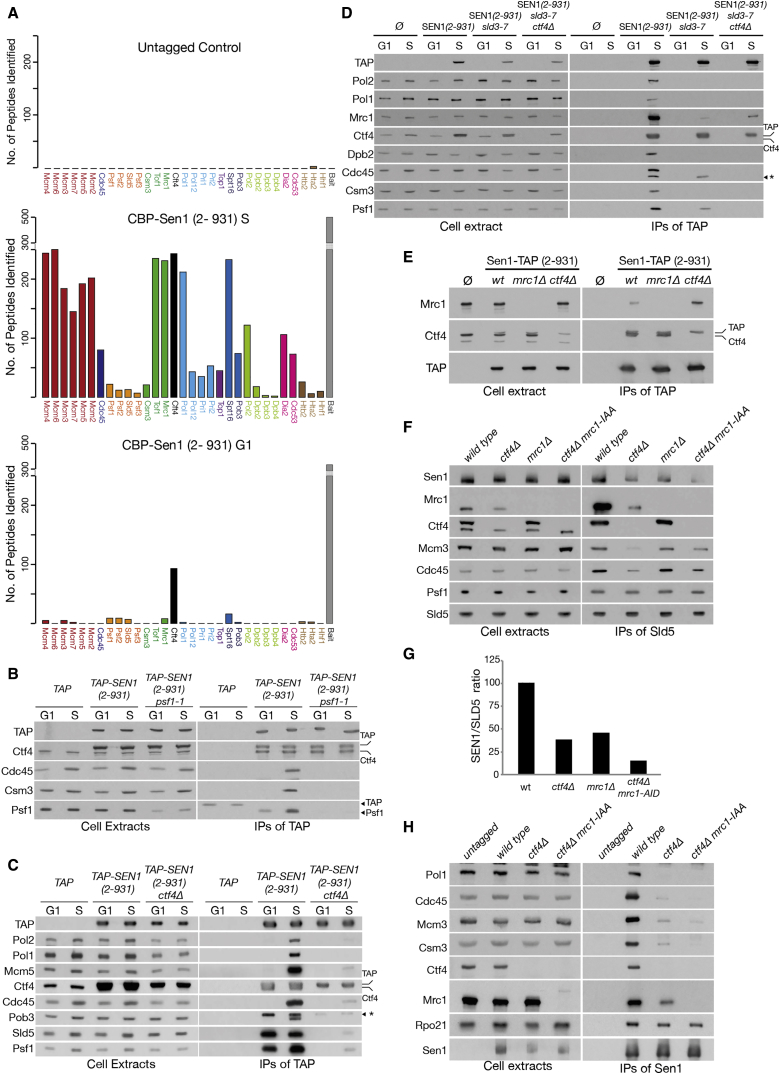
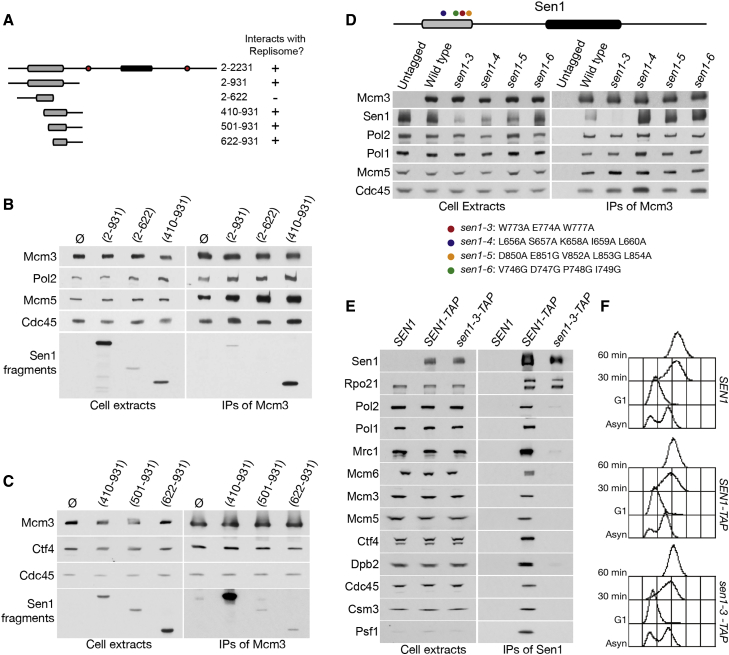
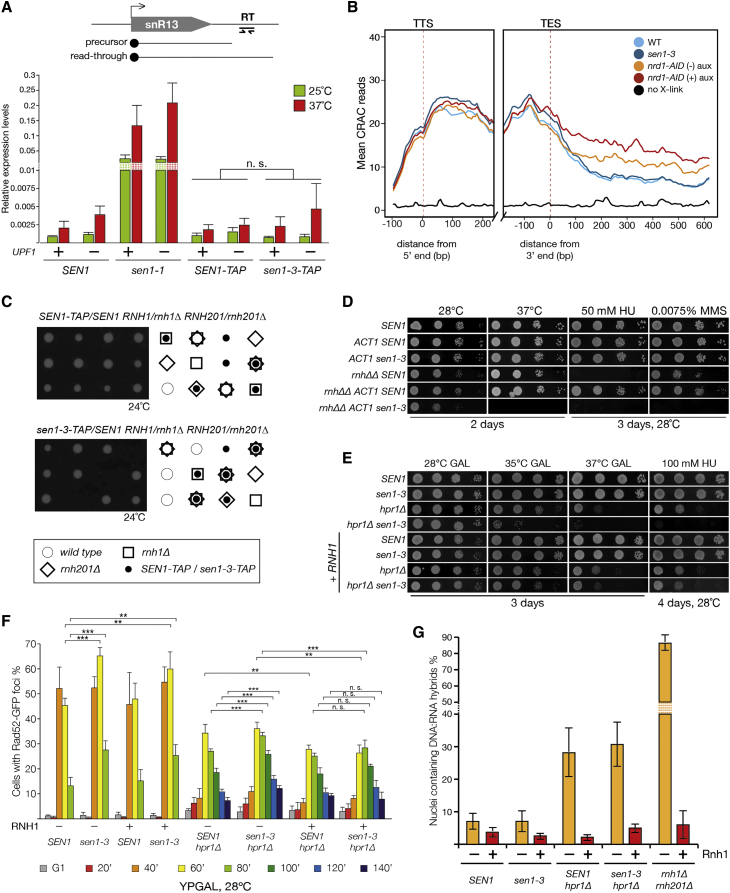
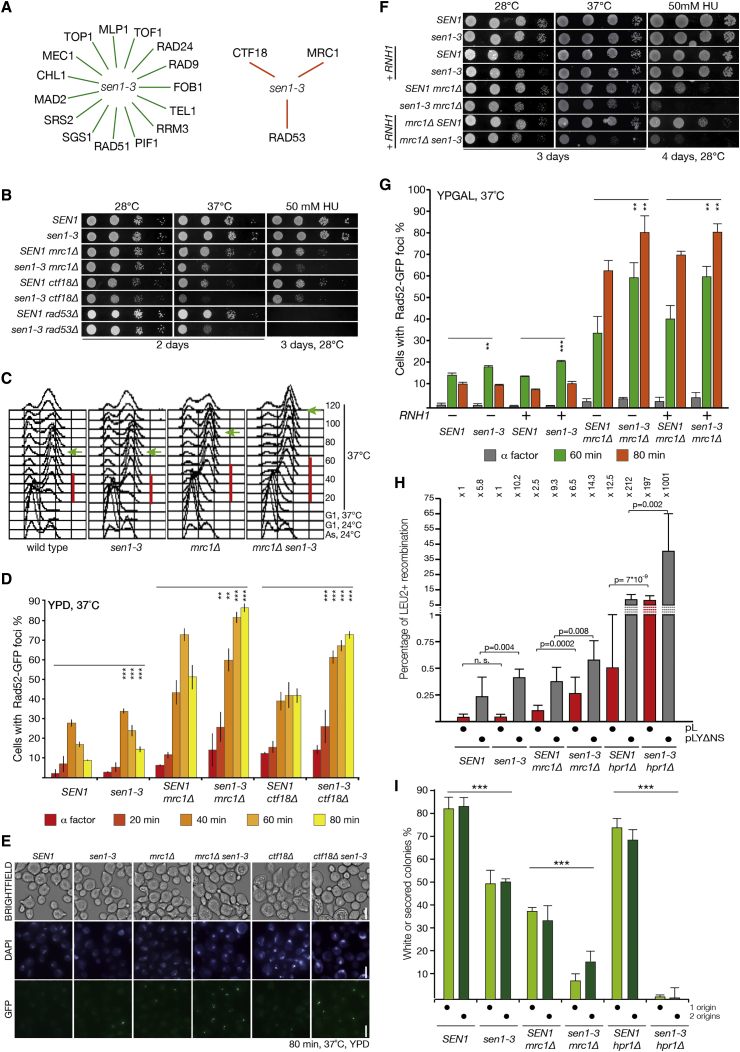
DNA replication and RNA transcription compete for the same substrate during S phase. Cells have evolved several mechanisms to minimize such conflicts. Here, we identify the mechanism by which the transcription termination helicase Sen1 associates with replisomes. We show that the N terminus of Sen1 is both sufficient and necessary for replisome association and that it binds to the replisome via the components Ctf4 and Mrc1. We generated a separation of function mutant, sen1-3, which abolishes replisome binding without affecting transcription termination. We observe that the sen1-3 mutants show increased genome instability and recombination levels. Moreover, sen1-3 is synthetically defective with mutations in genes involved in RNA metabolism and the S phase checkpoint. RNH1 overexpression suppresses defects in the former, but not the latter. These findings illustrate how Sen1 plays a key function at replication forks during DNA replication to promote fork progression and chromosome stability.
Keywords: Ctf4; DNA replication; DNA:RNA hybrids; Hpr1; Mrc1; RNA transcription; RNAse H; S phase checkpoint; Sen1; replisome.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Aguilera A., García-Muse T. R loops: from transcription byproducts to threats to genome stability. Mol. Cell. 2012;46:115–124. - PubMed
-
- Arigo J.T., Eyler D.E., Carroll K.L., Corden J.L. Termination of cryptic unstable transcripts is directed by yeast RNA-binding proteins Nrd1 and Nab3. Mol. Cell. 2006;23:841–851. - PubMed
-
- Bonnet A., Grosso A.R., Elkaoutari A., Coleno E., Presle A., Sridhara S.C., Janbon G., Géli V., de Almeida S.F., Palancade B. Introns protect eukaryotic genomes from transcription-associated genetic instability. Mol. Cell. 2017;67:608–621.e6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
