

Topography in the Bursting Dynamics of Entorhinal Neurons
- PMID: 32075768
- PMCID: PMC7053254
- DOI: 10.1016/j.celrep.2020.01.057
Topography in the Bursting Dynamics of Entorhinal Neurons
Abstract
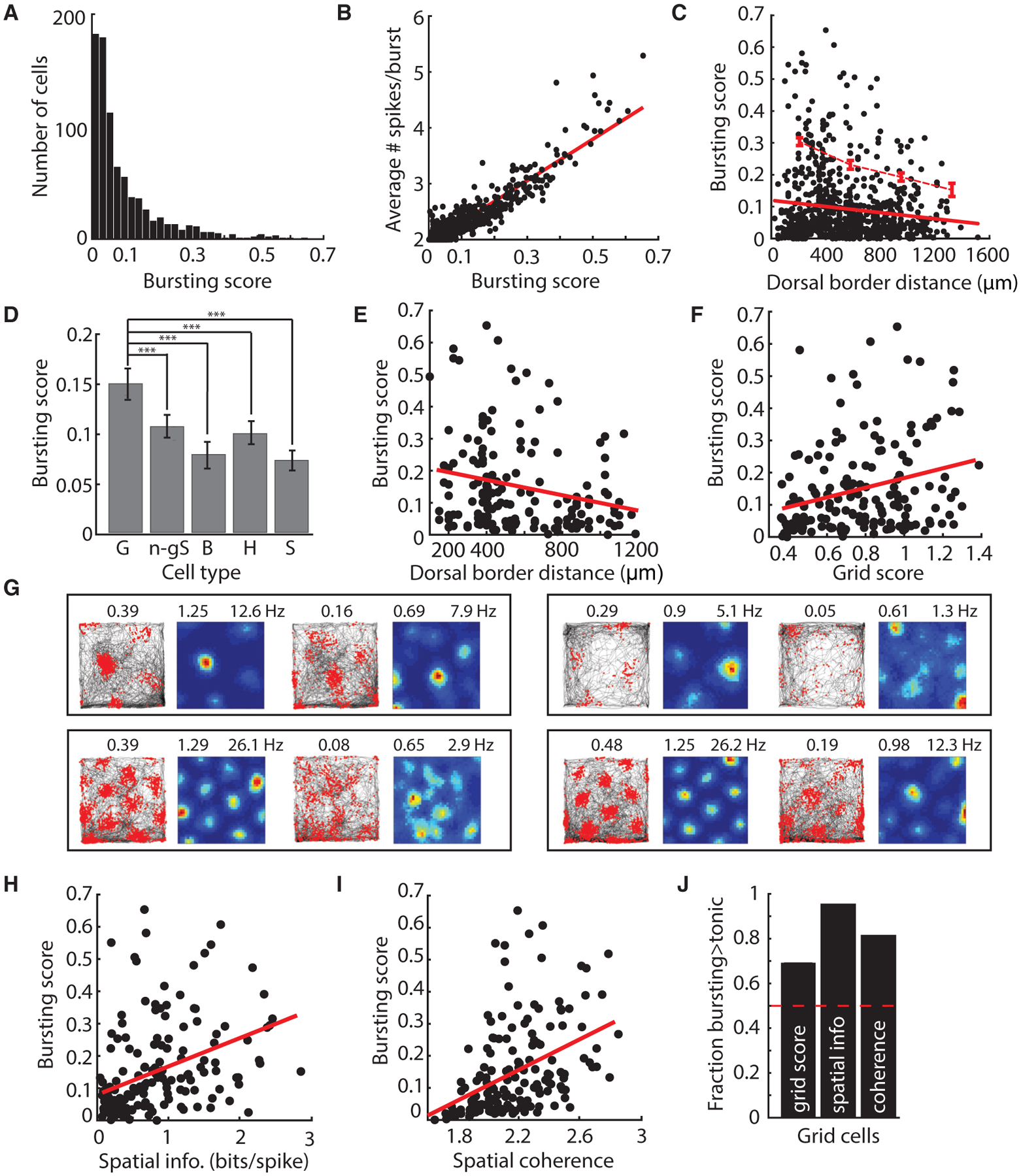
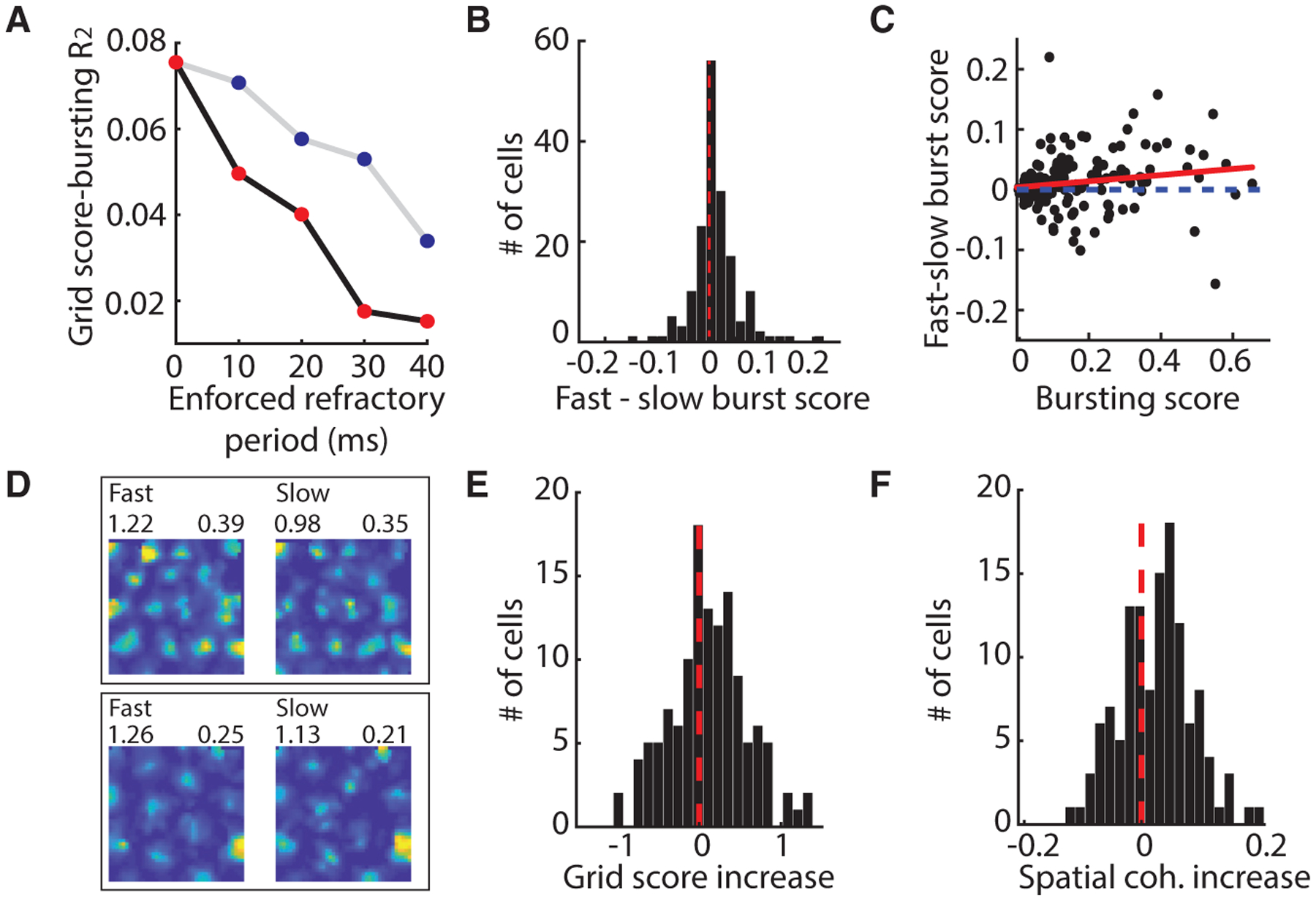
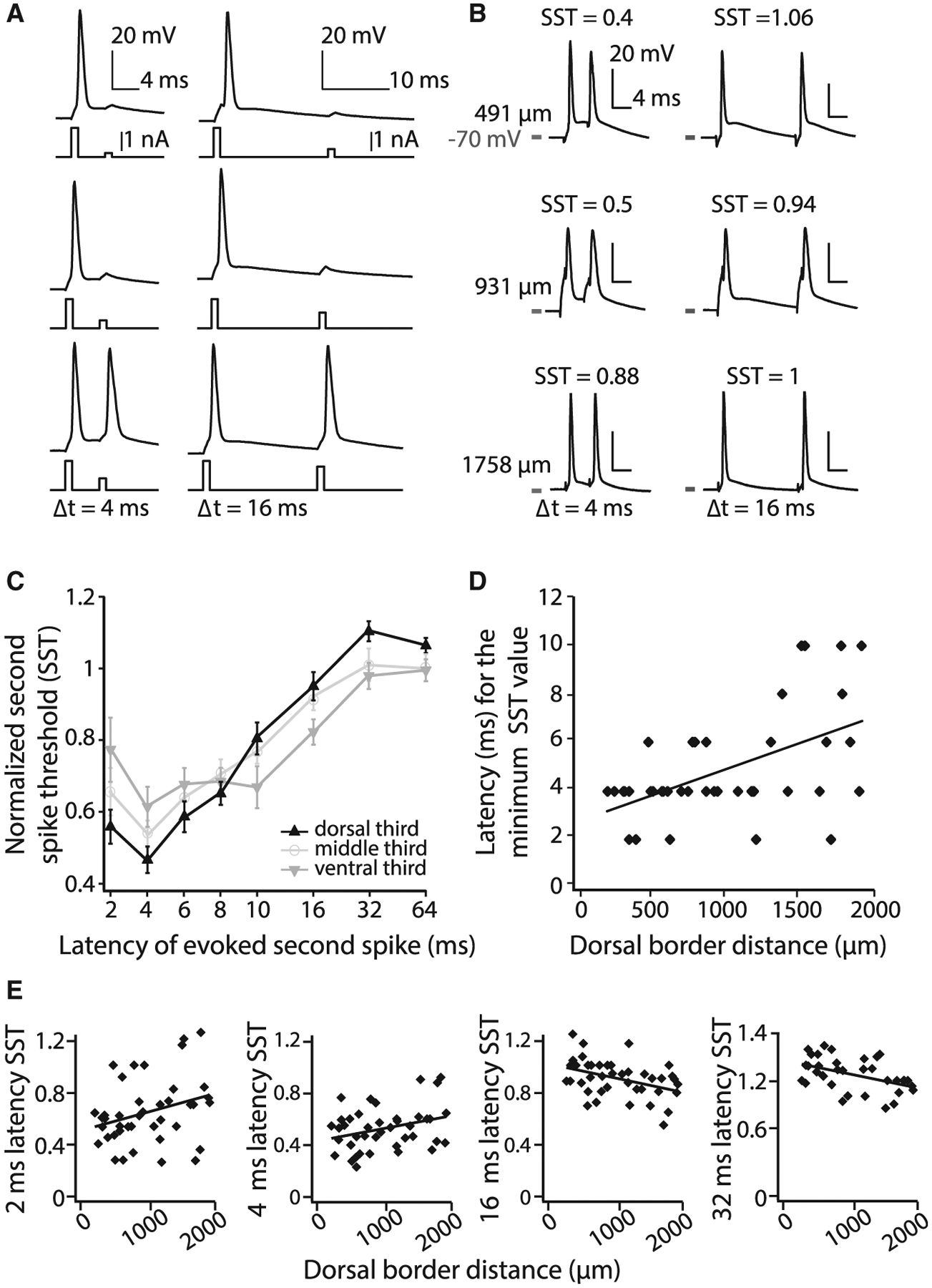
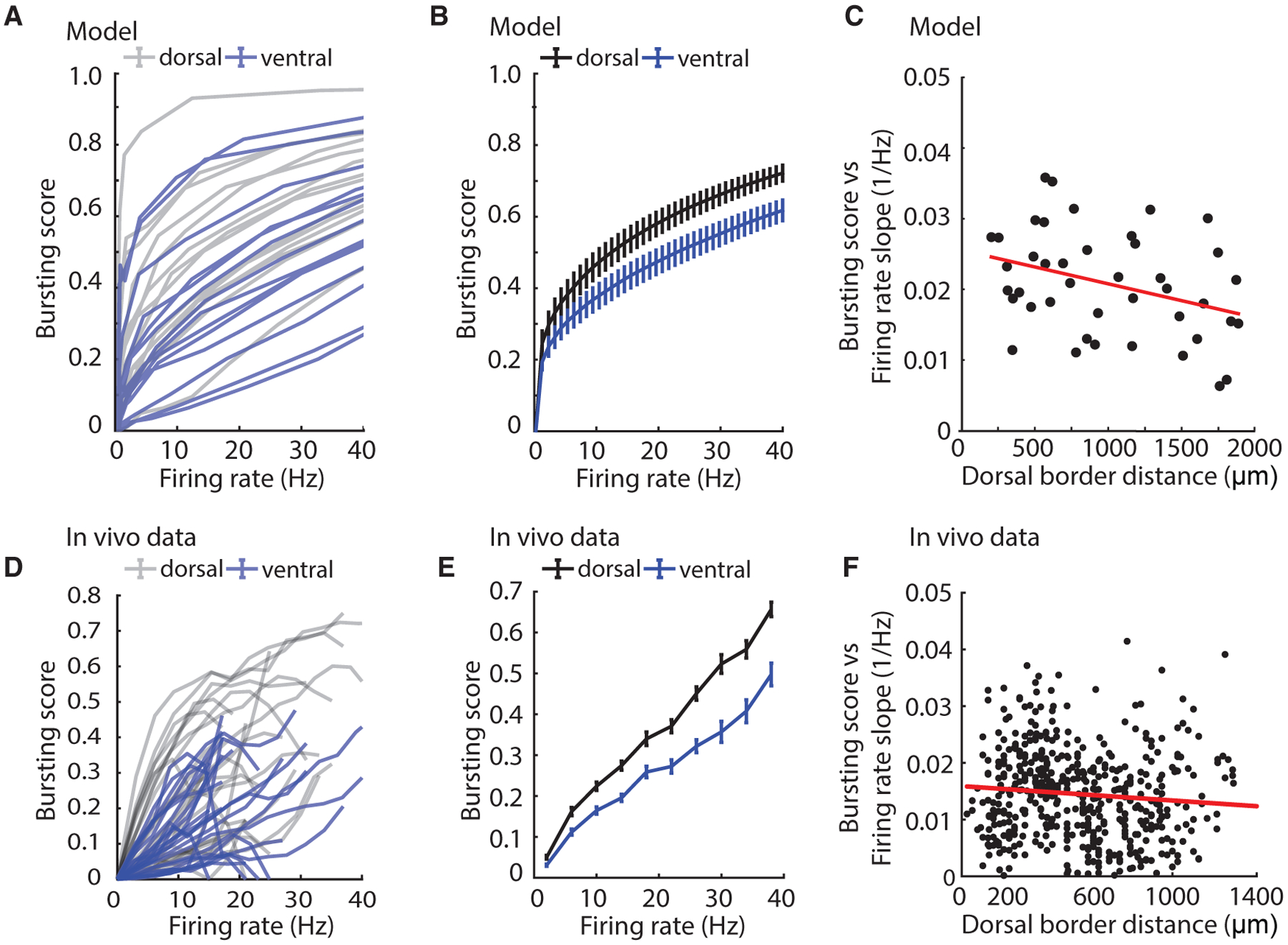
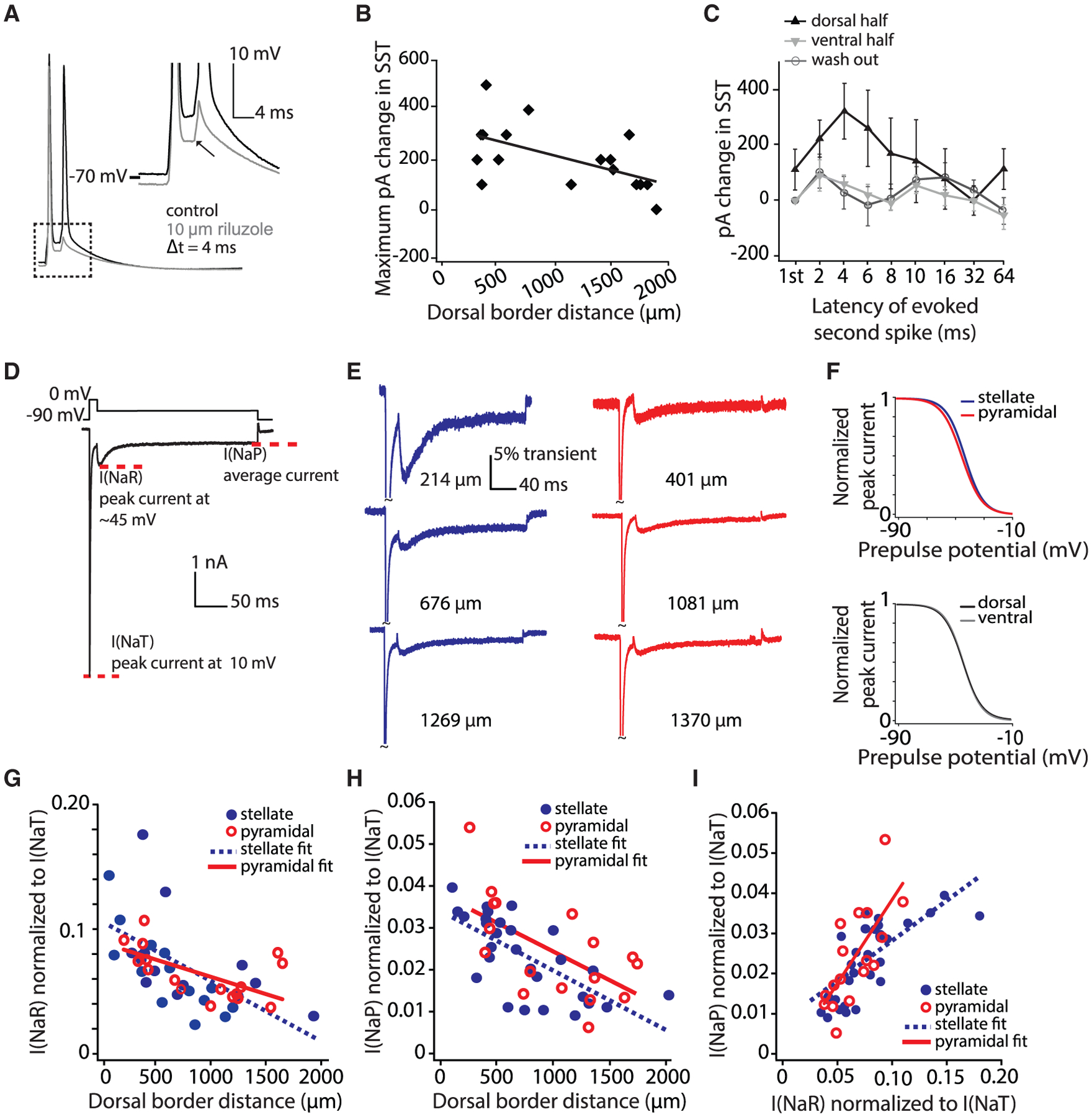
Medial entorhinal cortex contains neural substrates for representing space. These substrates include grid cells that fire in repeating locations and increase in scale progressively along the dorsal-to-ventral entorhinal axis, with the physical distance between grid firing nodes increasing from tens of centimeters to several meters in rodents. Whether the temporal scale of grid cell spiking dynamics shows a similar dorsal-to-ventral organization remains unknown. Here, we report the presence of a dorsal-to-ventral gradient in the temporal spiking dynamics of grid cells in behaving mice. This gradient in bursting supports the emergence of a dorsal grid cell population with a high signal-to-noise ratio. In vitro recordings combined with a computational model point to a role for gradients in non-inactivating sodium conductances in supporting the bursting gradient in vivo. Taken together, these results reveal a complementary organization in the temporal and intrinsic properties of entorhinal cells.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Alonso A, and Klink R (1993). Differential electroresponsiveness of stellate and pyramidal-like cells of medial entorhinal cortex layer II. J. Neurophysiol 70, 128–143. - PubMed
-
- Alonso A, and Llinás RR (1989). Subthreshold Na+-dependent theta-like rhythmicity in stellate cells of entorhinal cortex layer II. Nature 342, 175–177. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
