

Structural basis of specific inhibition of extracellular activation of pro- or latent myostatin by the monoclonal antibody SRK-015
- PMID: 32075906
- PMCID: PMC7170532
- DOI: 10.1074/jbc.RA119.012293
Structural basis of specific inhibition of extracellular activation of pro- or latent myostatin by the monoclonal antibody SRK-015
Abstract
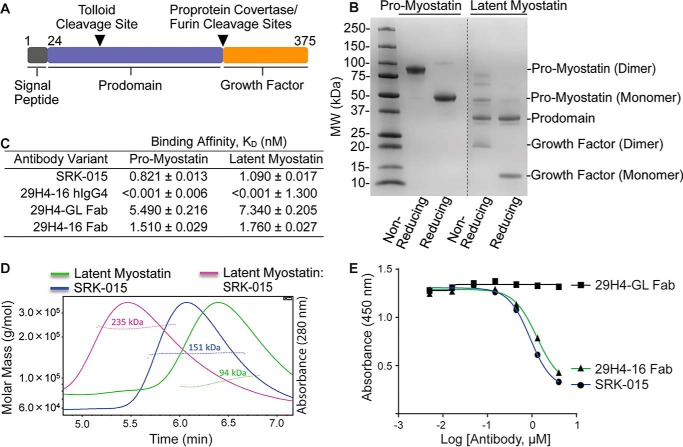
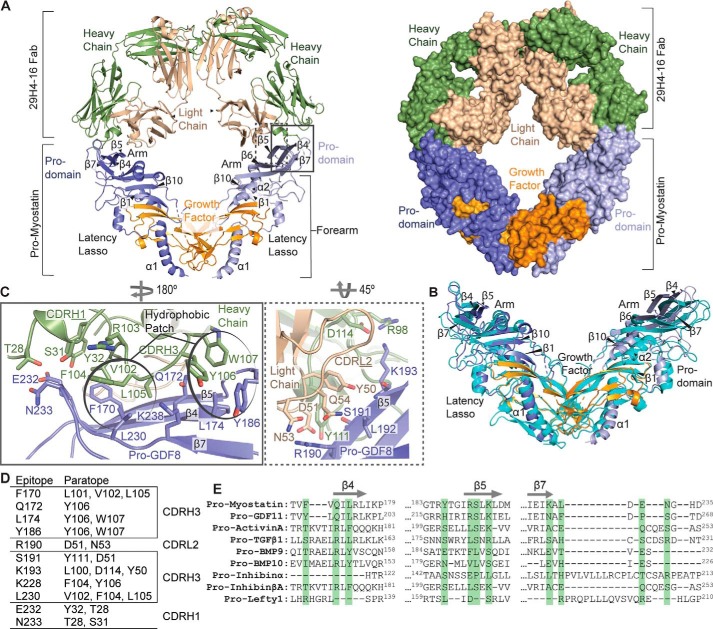
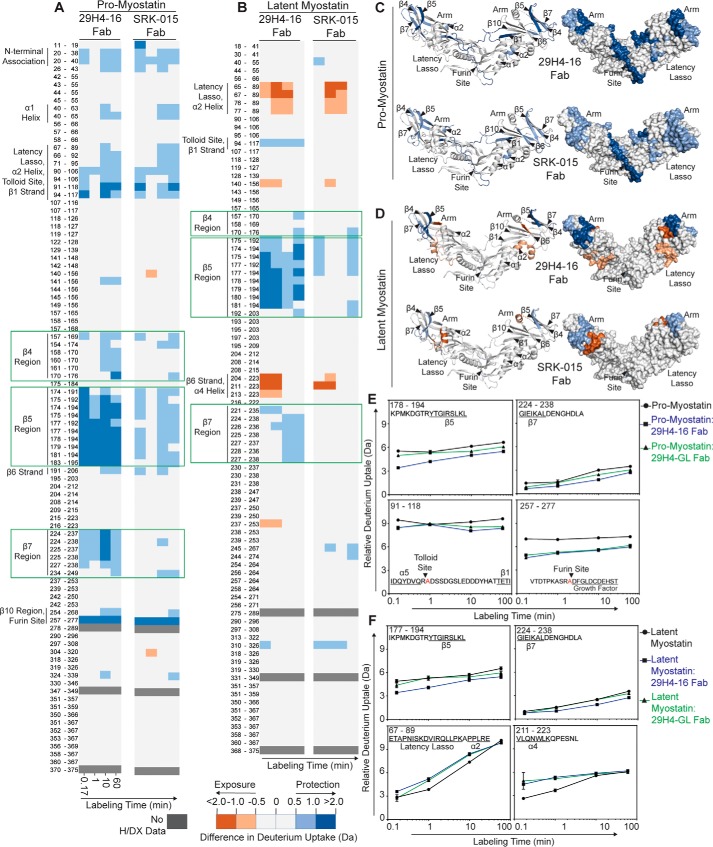
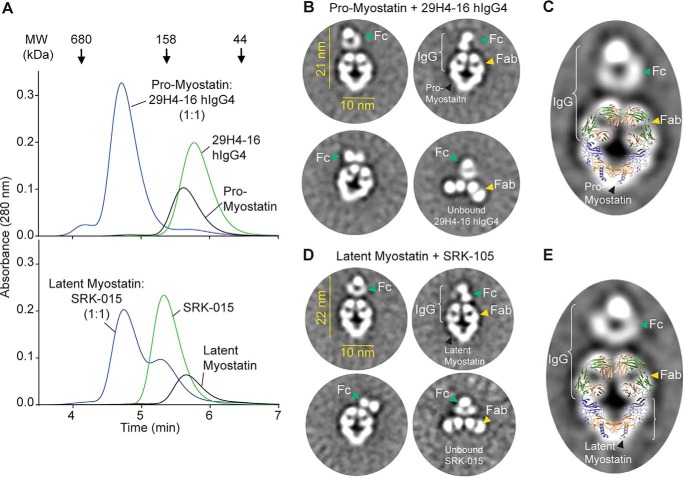
Myostatin (or growth/differentiation factor 8 (GDF8)) is a member of the transforming growth factor β superfamily of growth factors and negatively regulates skeletal muscle growth. Its dysregulation is implicated in muscle wasting diseases. SRK-015 is a clinical-stage mAb that prevents extracellular proteolytic activation of pro- and latent myostatin. Here we used integrated structural and biochemical approaches to elucidate the molecular mechanism of antibody-mediated neutralization of pro-myostatin activation. The crystal structure of pro-myostatin in complex with 29H4-16 Fab, a high-affinity variant of SRK-015, at 2.79 Å resolution revealed that the antibody binds to a conformational epitope in the arm region of the prodomain distant from the proteolytic cleavage sites. This epitope is highly sequence-divergent, having only limited similarity to other closely related members of the transforming growth factor β superfamily. Hydrogen/deuterium exchange MS experiments indicated that antibody binding induces conformational changes in pro- and latent myostatin that span the arm region, the loops contiguous to the protease cleavage sites, and the latency-associated structural elements. Moreover, negative-stain EM with full-length antibodies disclosed a stable, ring-like antigen-antibody structure in which the two Fab arms of a single antibody occupy the two arm regions of the prodomain in the pro- and latent myostatin homodimers, suggesting a 1:1 (antibody:myostatin homodimer) binding stoichiometry. These results suggest that SRK-015 binding stabilizes the latent conformation and limits the accessibility of protease cleavage sites within the prodomain. These findings shed light on approaches that specifically block the extracellular activation of growth factors by targeting their precursor forms.
Keywords: X-ray crystallography; cell signaling; growth differentiation factor 8 (GDF8); hydrogen exchange mass spectrometry; inhibition mechanism; monoclonal antibody; muscle atrophy; myostatin; proteolysis; skeletal muscle; transforming growth factor β (TGF-β).
© 2020 Dagbay et al.
Conflict of interest statement
All authors are current or former employees of Scholar Rock, Inc. and may be stock and/or stock option holders. G.C. and J.J. are named inventors on patents or patent applications related to SRK-015
Figures
References
-
- Reisz-Porszasz S., Bhasin S., Artaza J. N., Shen R., Sinha-Hikim I., Hogue A., Fielder T. J., and Gonzalez-Cadavid N. F. (2003) Lower skeletal muscle mass in male transgenic mice with muscle-specific overexpression of myostatin. Am. J. Physiol. Endocrinol. Metab. 285, E876–E888 10.1152/ajpendo.00107.2003 - DOI - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
