

Twenty Years of SynGAP Research: From Synapses to Cognition
- PMID: 32075947
- PMCID: PMC7046327
- DOI: 10.1523/JNEUROSCI.0420-19.2020
Twenty Years of SynGAP Research: From Synapses to Cognition
Abstract
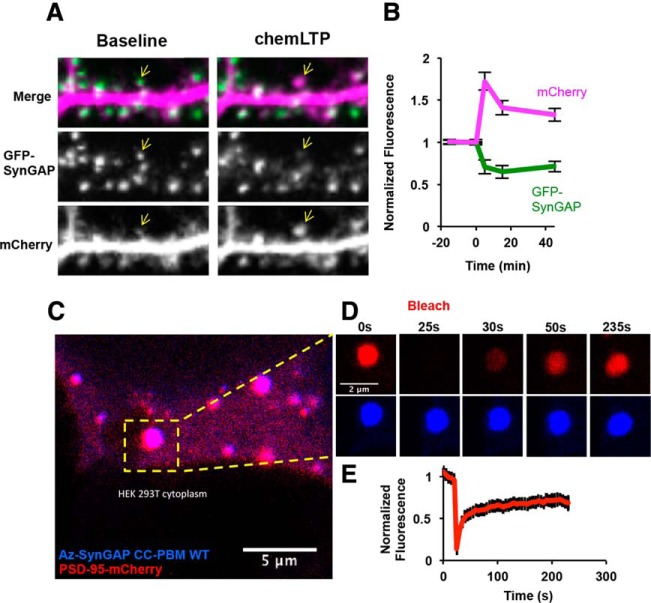
SynGAP is a potent regulator of biochemical signaling in neurons and plays critical roles in neuronal function. It was first identified in 1998, and has since been extensively characterized as a mediator of synaptic plasticity. Because of its involvement in synaptic plasticity, SynGAP has emerged as a critical protein for normal cognitive function. In recent years, mutations in the SYNGAP1 gene have been shown to cause intellectual disability in humans and have been linked to other neurodevelopmental disorders, such as autism spectrum disorders and schizophrenia. While the structure and biochemical function of SynGAP have been well characterized, a unified understanding of the various roles of SynGAP at the synapse and its contributions to neuronal function remains to be achieved. In this review, we summarize and discuss the current understanding of the multifactorial role of SynGAP in regulating neuronal function gathered over the last two decades.
Copyright © 2020 the authors.
Figures




References
-
- Aceti M, Creson TK, Vaissiere T, Rojas C, Huang WC, Wang YX, Petralia RS, Page DT, Miller CA, Rumbaugh G (2015) Syngap1 haploinsufficiency damages a postnatal critical period of pyramidal cell structural maturation linked to cortical circuit assembly. Biol Psychiatry 77:805–815. 10.1016/j.biopsych.2014.08.001 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources