

A phylogenetic model for the recruitment of species into microbial communities and application to studies of the human microbiome
- PMID: 32076128
- PMCID: PMC7242462
- DOI: 10.1038/s41396-020-0613-7
A phylogenetic model for the recruitment of species into microbial communities and application to studies of the human microbiome
Abstract
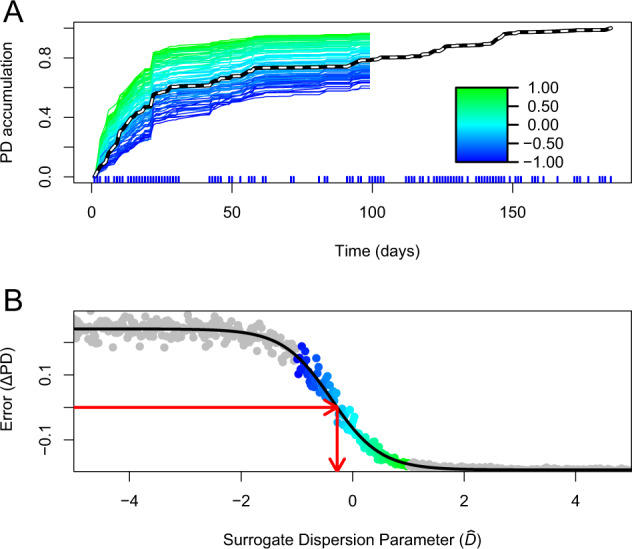
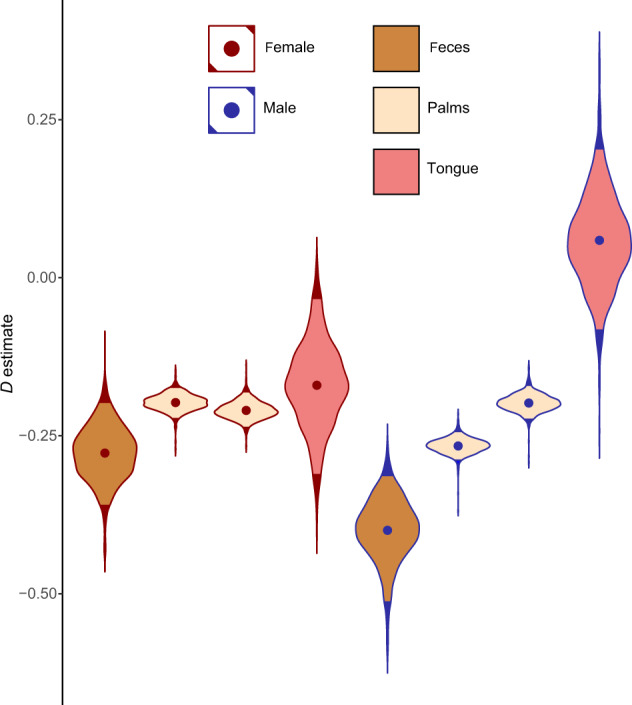
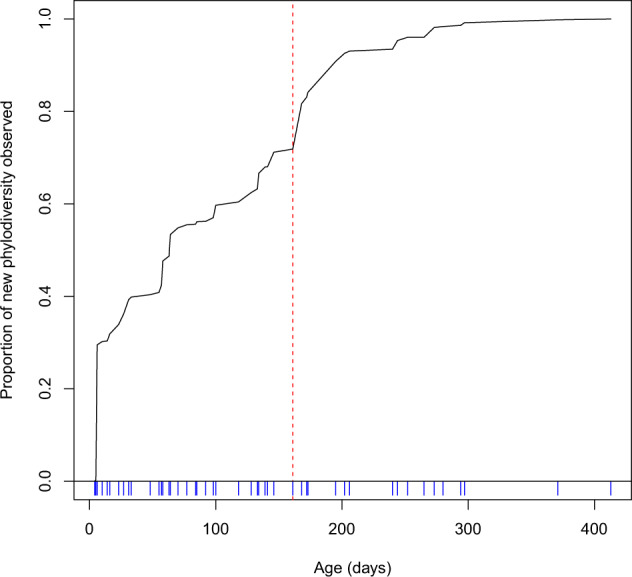
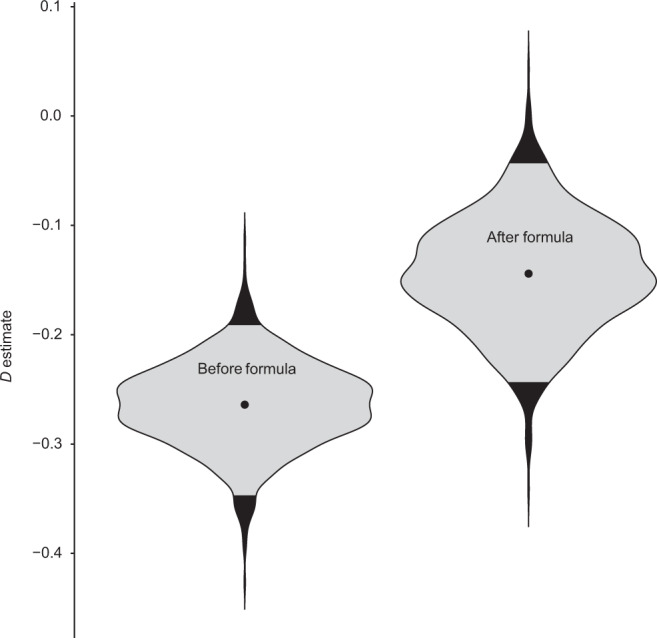
Understanding when and why new species are recruited into microbial communities is a formidable problem with implications for managing microbial systems, for instance by helping us better understand whether a probiotic or pathogen would be expected to colonize a human microbiome. Much theory in microbial temporal dynamics is focused on how phylogenetic relationships between microbes impact the order in which those microbes are recruited; for example, species that are closely related may competitively exclude each other. However, several recent human microbiome studies have observed closely related bacteria being recruited into microbial communities in short succession, suggesting that microbial community assembly is historically contingent, but competitive exclusion of close relatives may not be important. To address this, we developed a mathematical model that describes the order in which new species are detected in microbial communities over time within a phylogenetic framework. We use our model to test three hypothetical assembly modes: underdispersion (species recruitment is more likely if a close relative was previously detected), overdispersion (recruitment is more likely if a close relative has not been previously detected), and the neutral model (recruitment likelihood is not related to phylogenetic relationships among species). We applied our model to longitudinal human microbiome data, and found that for the individuals we analyzed, the human microbiome generally follows the underdispersion (i.e., nepotism) hypothesis. Exceptions were oral communities and the fecal communities of two infants that had undergone heavy antibiotic treatment. None of the datasets we analyzed showed statistically significant phylogenetic overdispersion.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Fukami T. Historical contingency in community assembly: integrating niches, species pools, and priority effects. Annu Rev Ecol Evol Syst. 2015;46:1–23.
