

Relaxed sequence constraints favor mutational freedom in idiosyncratic metazoan mitochondrial tRNAs
- PMID: 32080176
- PMCID: PMC7033119
- DOI: 10.1038/s41467-020-14725-y
Relaxed sequence constraints favor mutational freedom in idiosyncratic metazoan mitochondrial tRNAs
Abstract
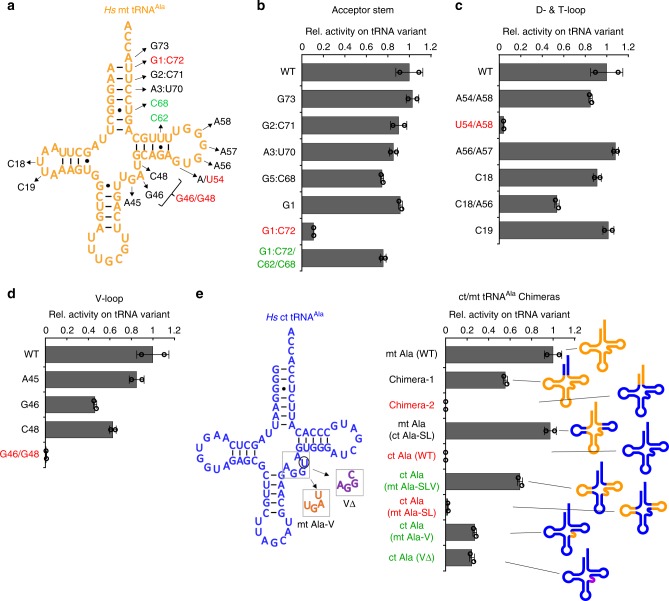
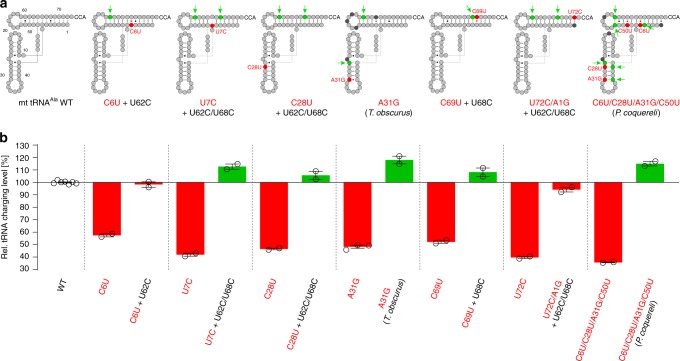
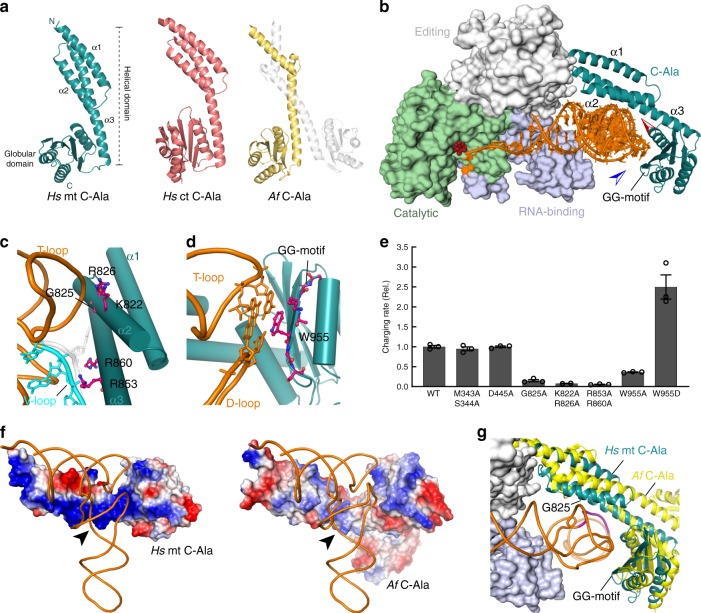
Metazoan complexity and life-style depend on the bioenergetic potential of mitochondria. However, higher aerobic activity and genetic drift impose strong mutation pressure and risk of irreversible fitness decline in mitochondrial (mt)DNA-encoded genes. Bilaterian mitochondria-encoded tRNA genes, key players in mitochondrial activity, have accumulated mutations at significantly higher rates than their cytoplasmic counterparts, resulting in foreshortened and fragile structures. Here we show that fragility of mt tRNAs coincided with the evolution of bilaterian animals. We demonstrate that bilaterians compensated for this reduced structural complexity in mt tRNAs by sequence-independent induced-fit adaption to the cognate mitochondrial aminoacyl-tRNA synthetase (aaRS). Structural readout by nuclear-encoded aaRS partners relaxed the sequence constraints on mt tRNAs and facilitated accommodation of functionally disruptive mutational insults by cis-acting epistatic compensations. Our results thus suggest that mutational freedom in mt tRNA genes is an adaptation to increased mutation pressure that was associated with the evolution of animal complexity.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Bains, W. in Evolutionary Biology (ed. Pontarotti, P.) 255–272 (Springer, Cham, 2016).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
