

Cardiac injury modulates critical components of prostaglandin E2 signaling during zebrafish heart regeneration
- PMID: 32080283
- PMCID: PMC7033201
- DOI: 10.1038/s41598-020-59868-6
Cardiac injury modulates critical components of prostaglandin E2 signaling during zebrafish heart regeneration
Abstract
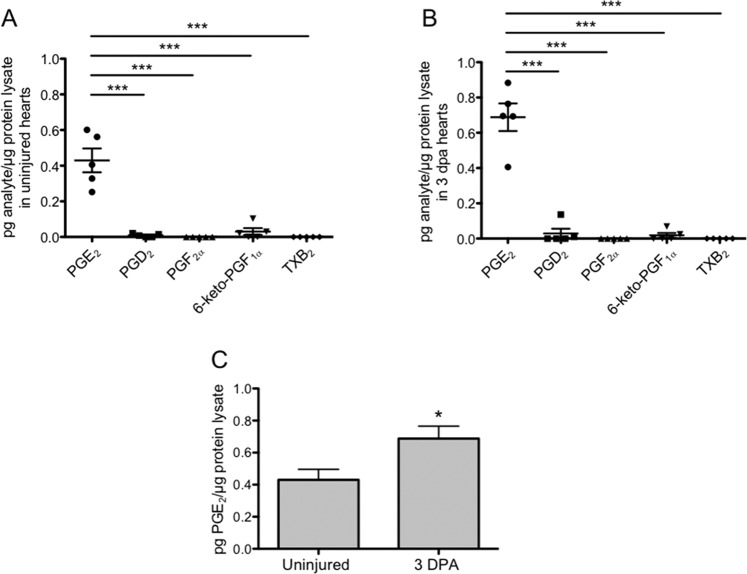
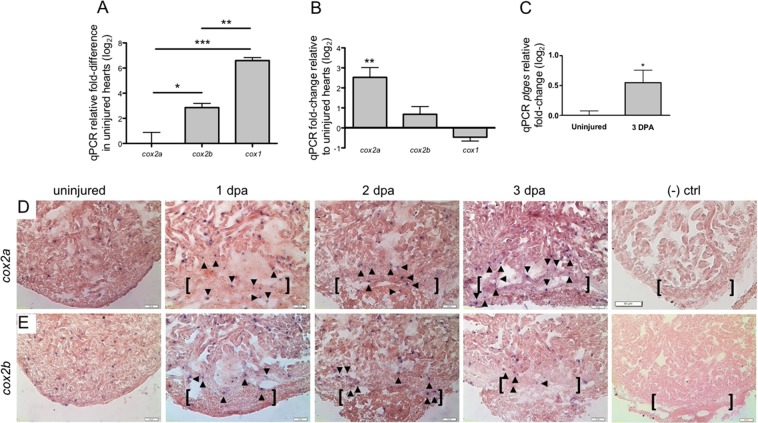
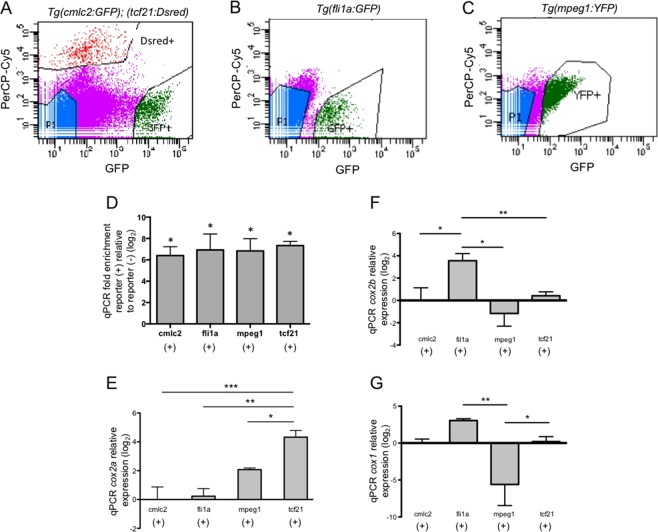
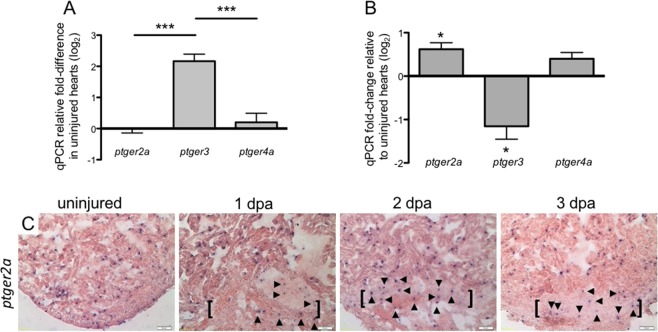
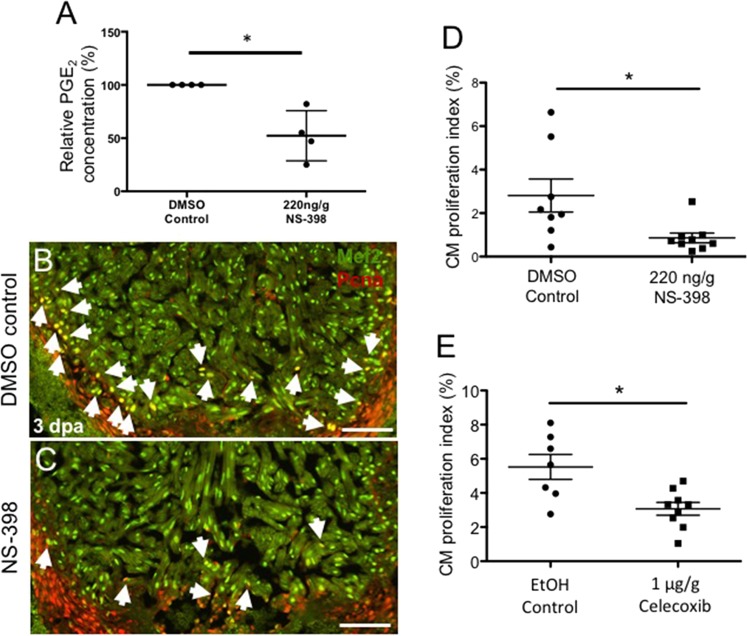
The inability to effectively stimulate cardiomyocyte proliferation remains a principle barrier to regeneration in the adult human heart. A tightly regulated, acute inflammatory response mediated by a range of cell types is required to initiate regenerative processes. Prostaglandin E2 (PGE2), a potent lipid signaling molecule induced by inflammation, has been shown to promote regeneration and cell proliferation; however, the dynamics of PGE2 signaling in the context of heart regeneration remain underexplored. Here, we employ the regeneration-competent zebrafish to characterize components of the PGE2 signaling circuit following cardiac injury. In the regenerating adult heart, we documented an increase in PGE2 levels, concurrent with upregulation of cox2a and ptges, two genes critical for PGE2 synthesis. Furthermore, we identified the epicardium as the most prominent site for cox2a expression, thereby suggesting a role for this tissue as an inflammatory mediator. Injury also drove the opposing expression of PGE2 receptors, upregulating pro-restorative ptger2a and downregulating the opposing receptor ptger3. Importantly, treatment with pharmacological inhibitors of Cox2 activity suppressed both production of PGE2, and the proliferation of cardiomyocytes. These results suggest that injury-induced PGE2 signaling is key to stimulating cardiomyocyte proliferation during regeneration.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Cano-Martinez A, et al. Functional and structural regeneration in the axolotl heart (Ambystoma mexicanum) after partial ventricular amputation. Arch. Cardiol. Mex. 2010;80:79–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
