

Antiviral activity of the natural alkaloid anisomycin against dengue and Zika viruses
- PMID: 32081740
- PMCID: PMC7279513
- DOI: 10.1016/j.antiviral.2020.104749
Antiviral activity of the natural alkaloid anisomycin against dengue and Zika viruses
Abstract
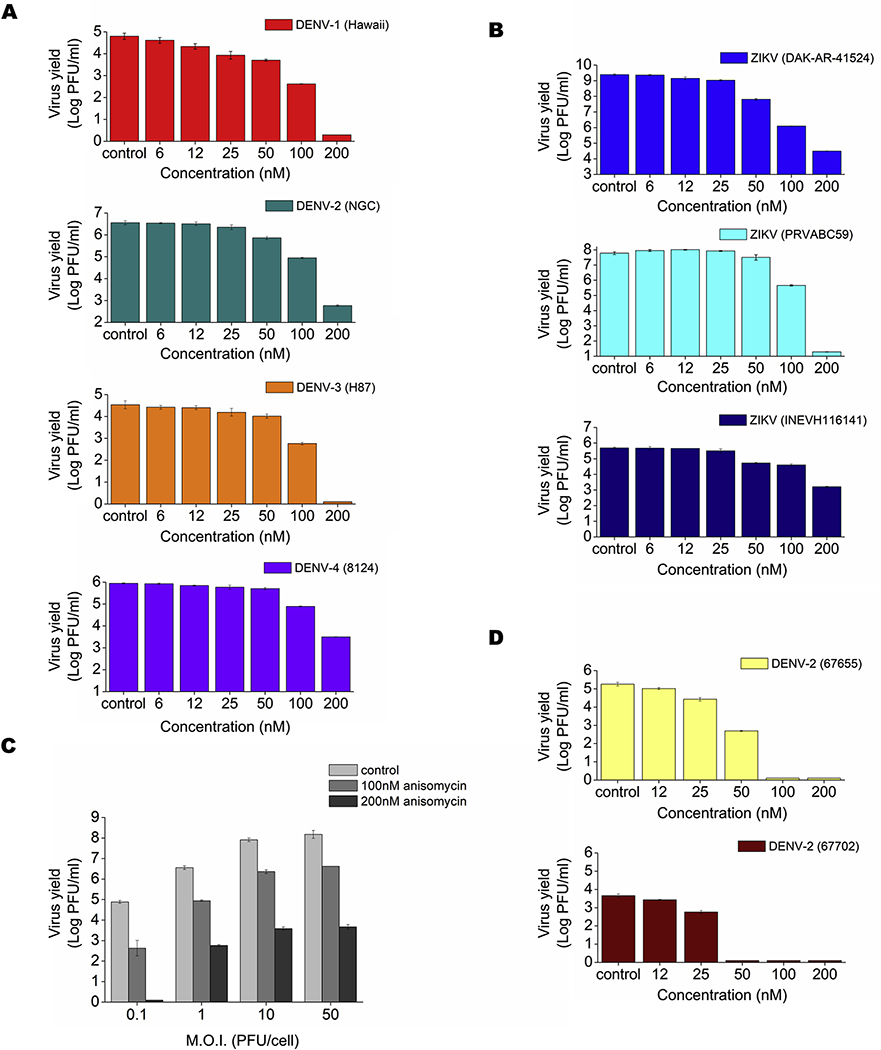
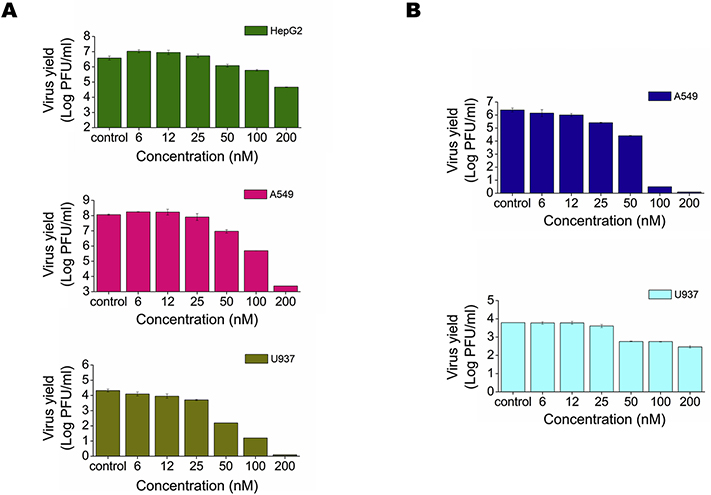
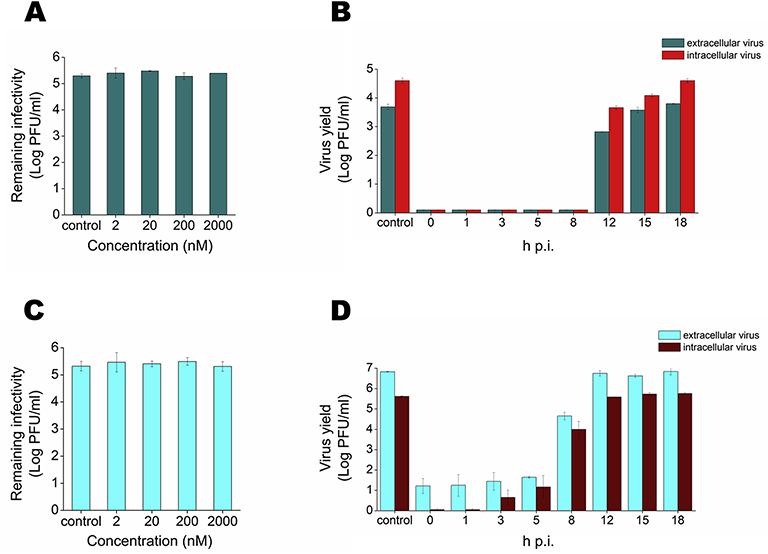
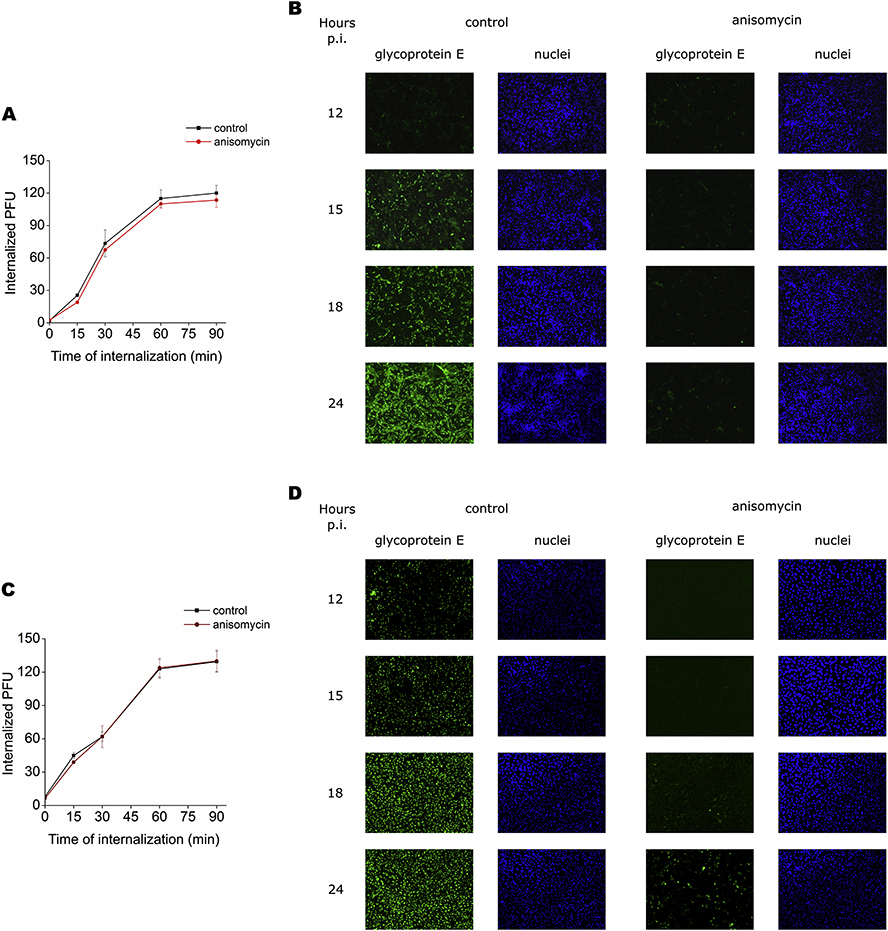
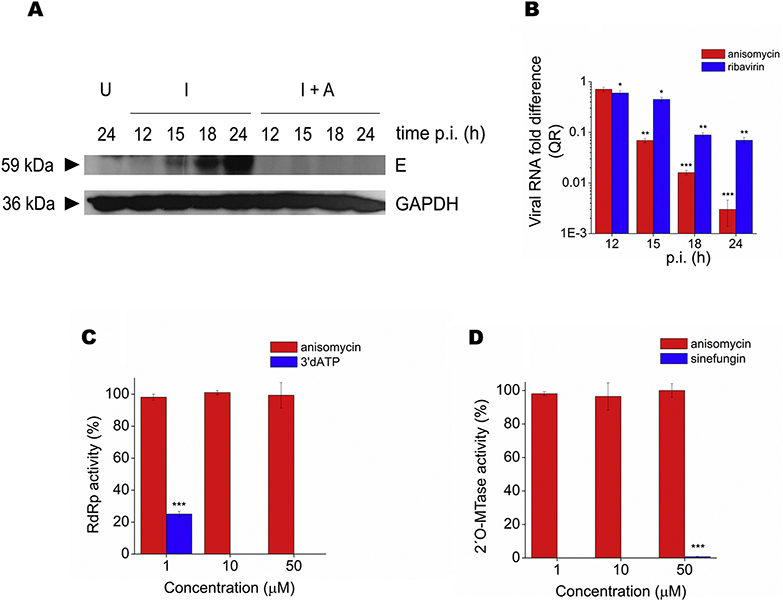
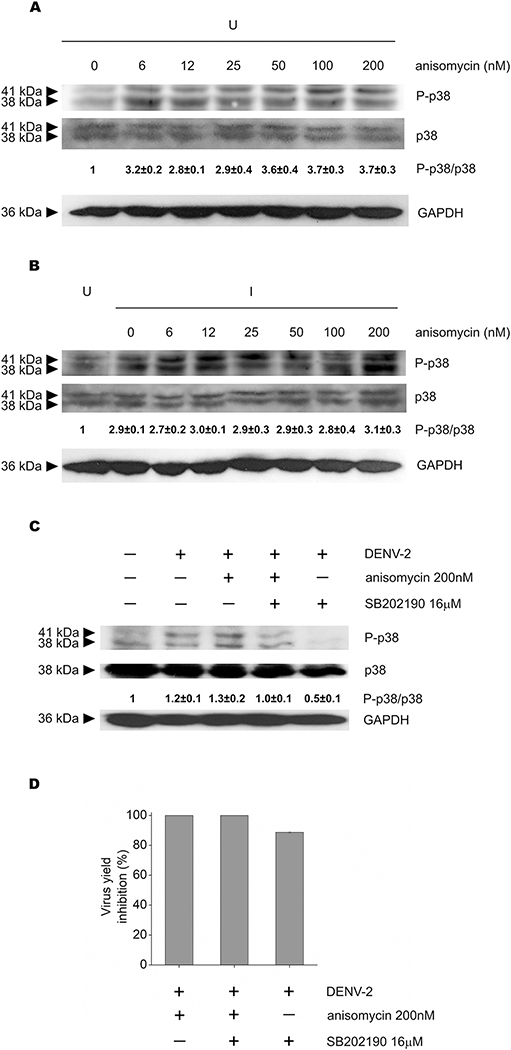
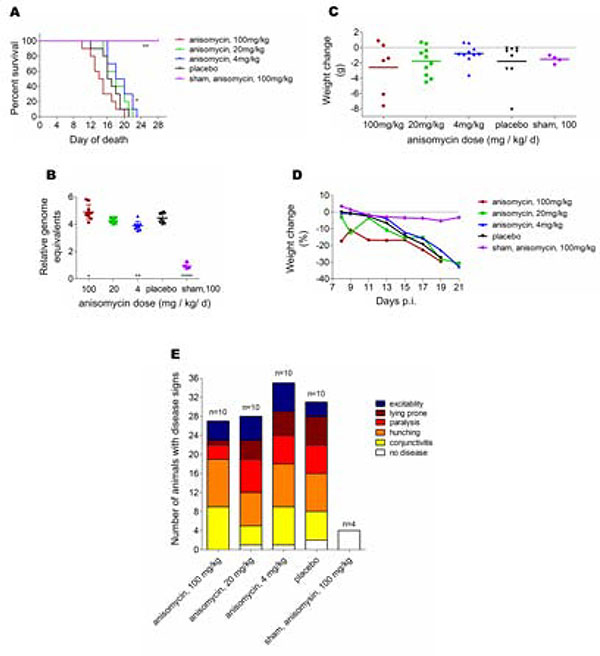
Flaviviruses constitute a public health concern because of their global burden and the lack of specific antiviral treatment. Here we investigated the antiviral activity of the alkaloid anisomycin against dengue (DENV) and Zika (ZIKV) viruses. At non-cytotoxic concentrations, anisomycin strongly inhibited the replication of reference strains and clinical isolates of all DENV serotypes and Asian and African strains of ZIKV in Vero cells. Anisomycin also prevented DENV and ZIKV multiplication in human cell lines. While initial steps of DENV and ZIKV replicative cycle were unaffected, a high inhibition of viral protein expression was demonstrated after treatment with anisomycin. DENV RNA synthesis was strongly reduced in anisomycin treated cultures, but the compound did not exert a direct inhibitory effect on 2' O-methyltransferase or RNA polymerase activities of DENV NS5 protein. Furthermore, anisomycin-mediated activation of p38 signaling was not related to the antiviral action of the compound. The evaluation of anisomycin efficacy in a mouse model of ZIKV morbidity and mortality revealed that animals treated with a low dose of anisomycin exhibited a significant reduction in viremia levels and died significantly later than the control group. This protective effect was lost at higher doses, though. In conclusion, anisomycin is a potent and selective in vitro inhibitor of DENV and ZIKV that impairs a post-entry step of viral replication; and a low-dose anisomycin treatment may provide some minimal benefit in a mouse model.
Keywords: Anisomycin; Antiviral; Dengue; Flavivirus; Zika; p38.
Copyright © 2020 Elsevier B.V. All rights reserved.
Figures
References
-
- Baud D, Gubler DJ, Schaub B, Lanteri MC, Musso D. An update on Zika virus infection. Lancet. 2017; 390(10107):2099–2109. - PubMed
-
- Benmansour F, Eydoux C, Querat G, de Lamballerie X, Canard B, Alvarez K, Guillemot JC, Novel Barral K. 2-phenyl-5-[(E)-2-(thiophen-2-yl)ethenyl]-1,3,4-oxadiazole and 3-phenyl-5-[(E)-2-(thiophen-2-yl)ethenyl]-1,2,4-oxadiazole derivatives as dengue virus inhibitors targeting NS5 polymerase. Eur J Med Chem. 2016; 109:146–56. - PubMed
-
- Ceballos-Olvera I, Chávez-Salinas S, Medina F, Ludert JE, del Angel RM. JNK phosphorylation, induced during dengue virus infection, is important for viral infection and requires the presence of cholesterol. Virology. 2010; 396(1):30–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
