

Systematic analysis of the Hippo pathway organization and oncogenic alteration in evolution
- PMID: 32081887
- PMCID: PMC7035326
- DOI: 10.1038/s41598-020-60120-4
Systematic analysis of the Hippo pathway organization and oncogenic alteration in evolution
Abstract
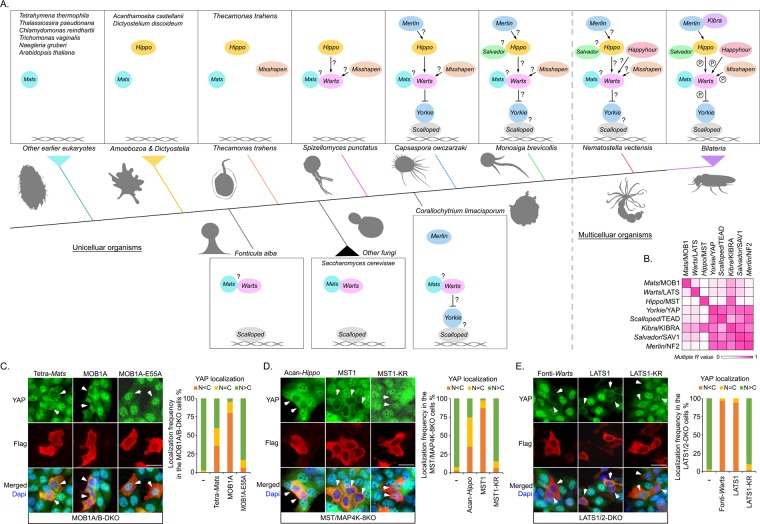
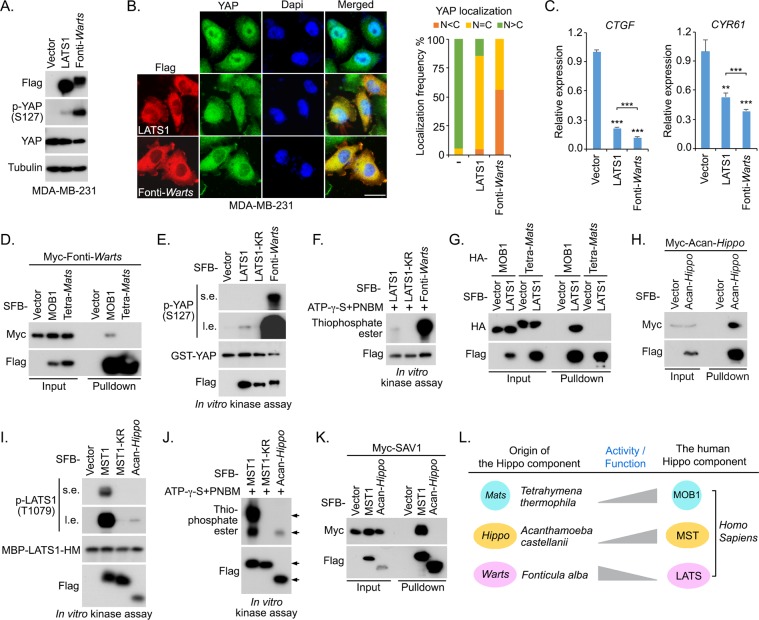
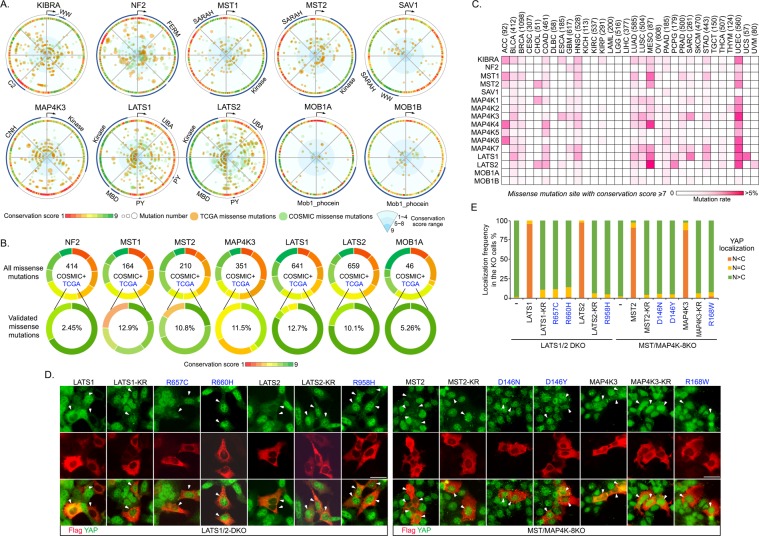
The Hippo pathway is a central regulator of organ size and a key tumor suppressor via coordinating cell proliferation and death. Initially discovered in Drosophila, the Hippo pathway has been implicated as an evolutionarily conserved pathway in mammals; however, how this pathway was evolved to be functional from its origin is still largely unknown. In this study, we traced the Hippo pathway in premetazoan species, characterized the intrinsic functions of its ancestor components, and unveiled the evolutionary history of this key signaling pathway from its unicellular origin. In addition, we elucidated the paralogous gene history for the mammalian Hippo pathway components and characterized their cancer-derived somatic mutations from an evolutionary perspective. Taken together, our findings not only traced the conserved function of the Hippo pathway to its unicellular ancestor components, but also provided novel evolutionary insights into the Hippo pathway organization and oncogenic alteration.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
