

The acid-base-nucleophile catalytic triad in ABH-fold enzymes is coordinated by a set of structural elements
- PMID: 32084230
- PMCID: PMC7034887
- DOI: 10.1371/journal.pone.0229376
The acid-base-nucleophile catalytic triad in ABH-fold enzymes is coordinated by a set of structural elements
Abstract
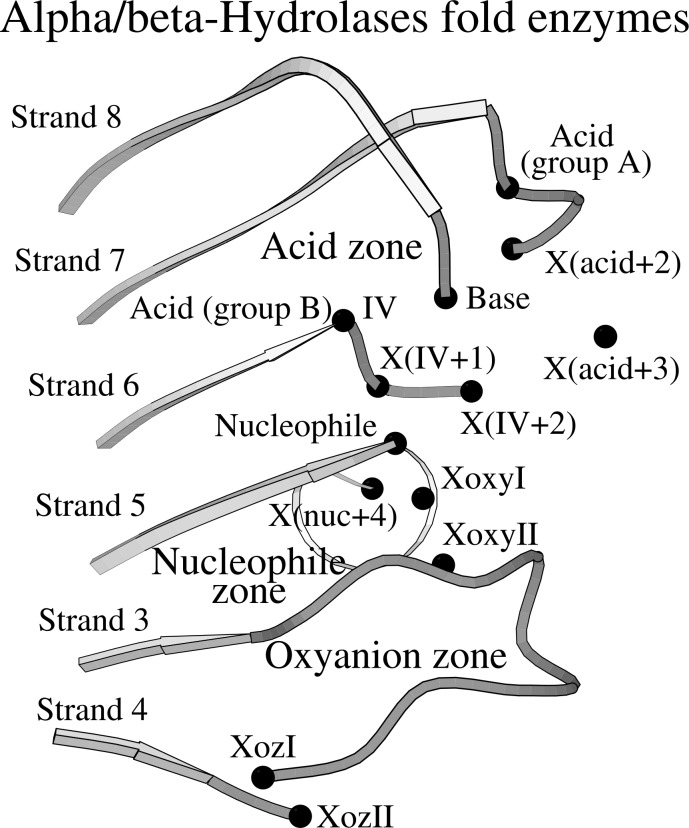
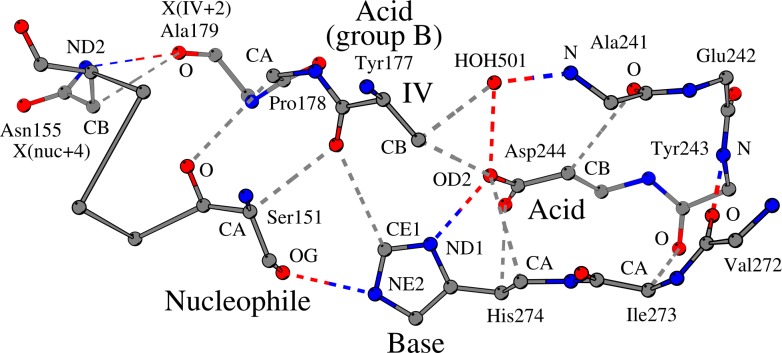
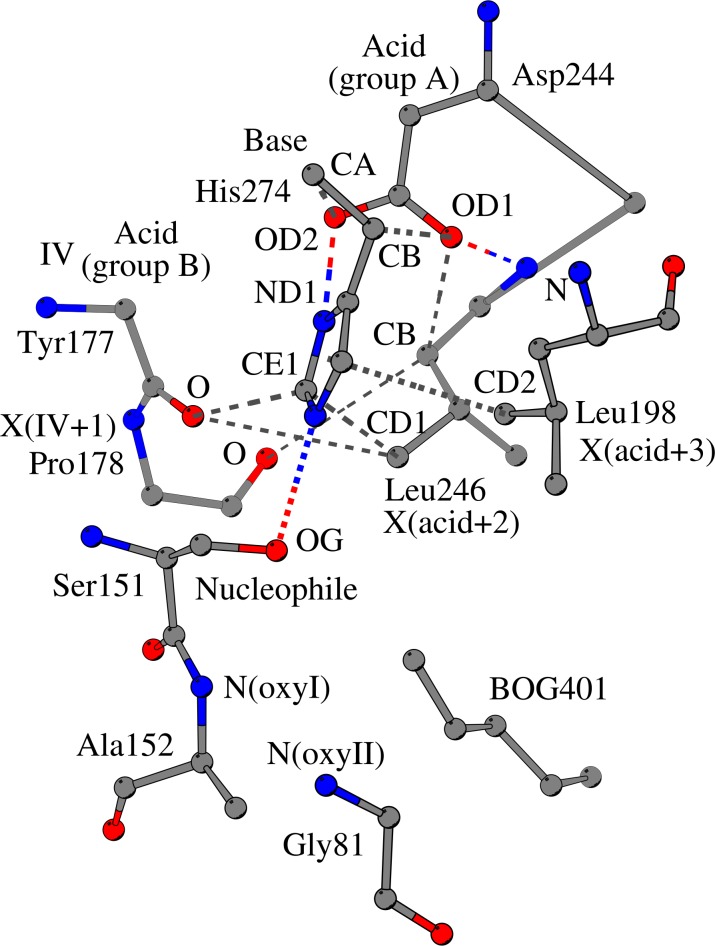
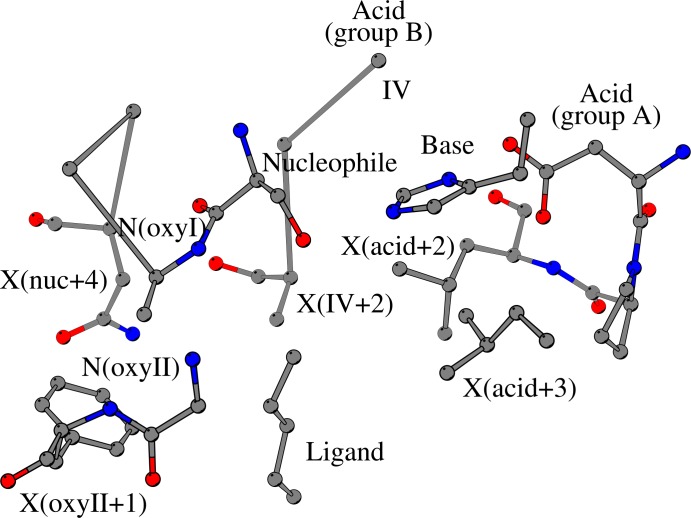
The alpha/beta-Hydrolases (ABH) are a structural class of proteins that are found widespread in nature and includes enzymes that can catalyze various reactions in different substrates. The catalytic versatility of the ABH fold enzymes, which has been a valuable property in protein engineering applications, is based on a similar acid-base-nucleophile catalytic mechanism. In our research, we are concerned with the structure that surrounds the key units of the catalytic machinery, and we have previously found conserved structural organizations that coordinate the catalytic acid, the catalytic nucleophile and the residues of the oxyanion hole. Here, we explore the architecture that surrounds the catalytic histidine at the active sites of enzymes from 40 ABH fold families, where we have identified six conserved interactions that coordinate the catalytic histidine next to the catalytic acid and the catalytic nucleophile. Specifically, the catalytic nucleophile is coordinated next to the catalytic histidine by two weak hydrogen bonds, while the catalytic acid is directly involved in the coordination of the catalytic histidine through by two weak hydrogen bonds. The imidazole ring of the catalytic histidine is coordinated by a CH-π contact and a hydrophobic interaction. Moreover, the catalytic triad residues are connected with a residue that is located at the core of the active site of ABH fold, which is suggested to be the fourth member of a "structural catalytic tetrad". Besides their role in the stability of the catalytic mechanism, the conserved elements of the catalytic site are actively involved in ligand binding and affect other properties of the catalytic activity, such as substrate specificity, enantioselectivity, pH optimum and thermostability of ABH fold enzymes. These properties are regularly targeted in protein engineering applications, and thus, the identified conserved structural elements can serve as potential modification sites in order to develop ABH fold enzymes with altered activities.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
