

A Metabolic Pathway for Activation of Dietary Glucosinolates by a Human Gut Symbiont
- PMID: 32084341
- PMCID: PMC7515767
- DOI: 10.1016/j.cell.2020.01.023
A Metabolic Pathway for Activation of Dietary Glucosinolates by a Human Gut Symbiont
Abstract
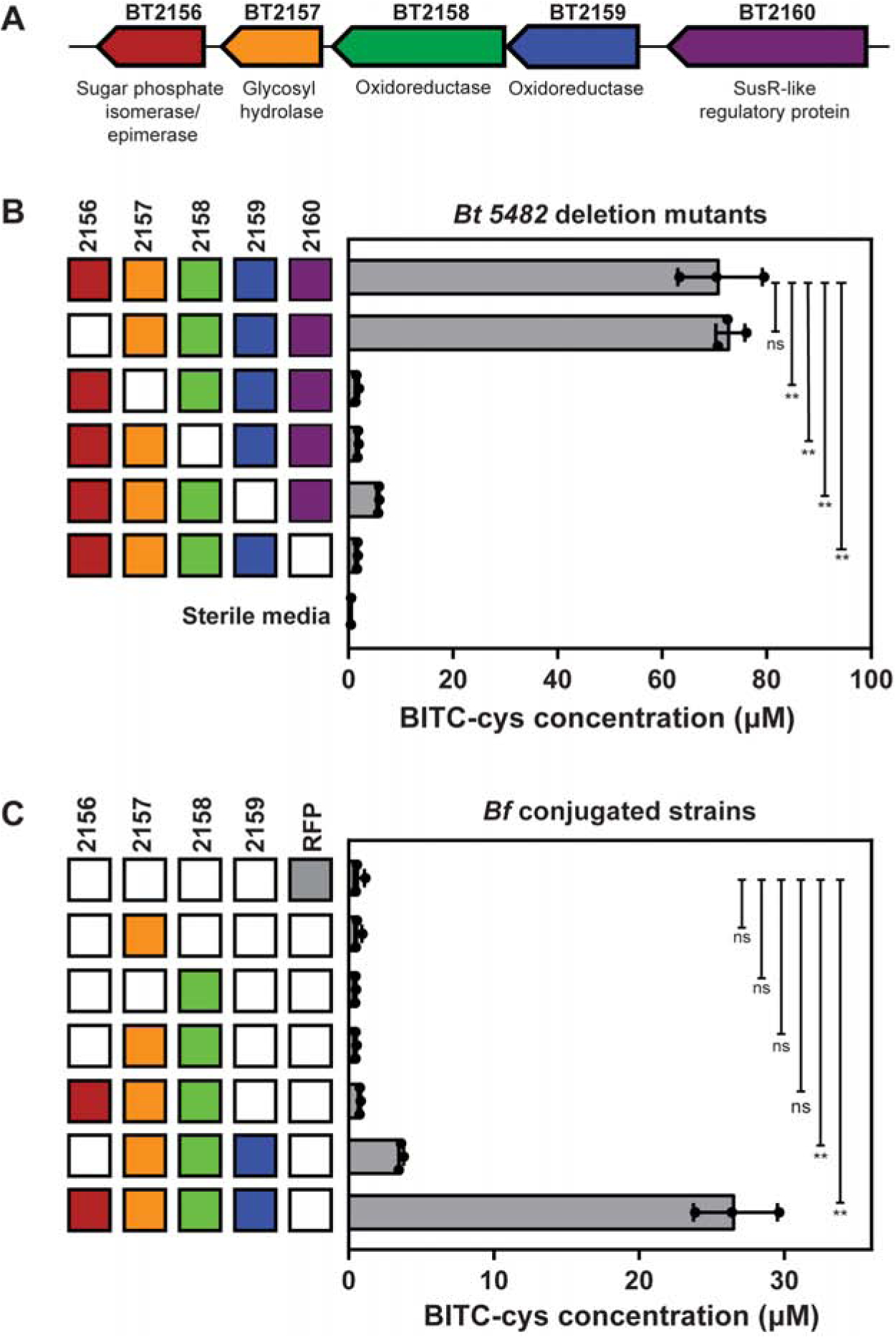
Consumption of glucosinolates, pro-drug-like metabolites abundant in Brassica vegetables, has been associated with decreased risk of certain cancers. Gut microbiota have the ability to metabolize glucosinolates, generating chemopreventive isothiocyanates. Here, we identify a genetic and biochemical basis for activation of glucosinolates to isothiocyanates by Bacteroides thetaiotaomicron, a prominent gut commensal species. Using a genome-wide transposon insertion screen, we identified an operon required for glucosinolate metabolism in B. thetaiotaomicron. Expression of BT2159-BT2156 in a non-metabolizing relative, Bacteroides fragilis, resulted in gain of glucosinolate metabolism. We show that isothiocyanate formation requires the action of BT2158 and either BT2156 or BT2157 in vitro. Monocolonization of mice with mutant BtΔ2157 showed reduced isothiocyanate production in the gastrointestinal tract. These data provide insight into the mechanisms by which a common gut bacterium processes an important dietary nutrient.
Keywords: Bacteroides thetaiotaomicron; glucosinolate; gut microbe; isothiocyanate; myrosinase; phytonutrient.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures




Comment in
-
Brassica vegetable metabolism by gut microbiota.Nat Rev Gastroenterol Hepatol. 2020 Apr;17(4):195. doi: 10.1038/s41575-020-0287-7. Nat Rev Gastroenterol Hepatol. 2020. PMID: 32161376 No abstract available.
References
-
- Agerbirk N, De Vos M, Kim JH, and Jander G (2009). Indole glucosinolate breakdown and its biological effects. Phytochem. Rev 8, 101–120.
-
- Albaser A, Kazana E, Bennett MH, Cebeci F, Luang-In V, Spanu PD, and Rossiter JT (2016). Discovery of a Bacterial Glycoside Hydrolase Family 3 (GH3) β-Glucosidase with Myrosinase Activity from a Citrobacter Strain Isolated from Soil. J. Agric. Food Chem 64, 1520–1527. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
