

Knocking out C9ORF72 Exacerbates Axonal Trafficking Defects Associated with Hexanucleotide Repeat Expansion and Reduces Levels of Heat Shock Proteins
- PMID: 32084385
- PMCID: PMC7066330
- DOI: 10.1016/j.stemcr.2020.01.010
Knocking out C9ORF72 Exacerbates Axonal Trafficking Defects Associated with Hexanucleotide Repeat Expansion and Reduces Levels of Heat Shock Proteins
Abstract
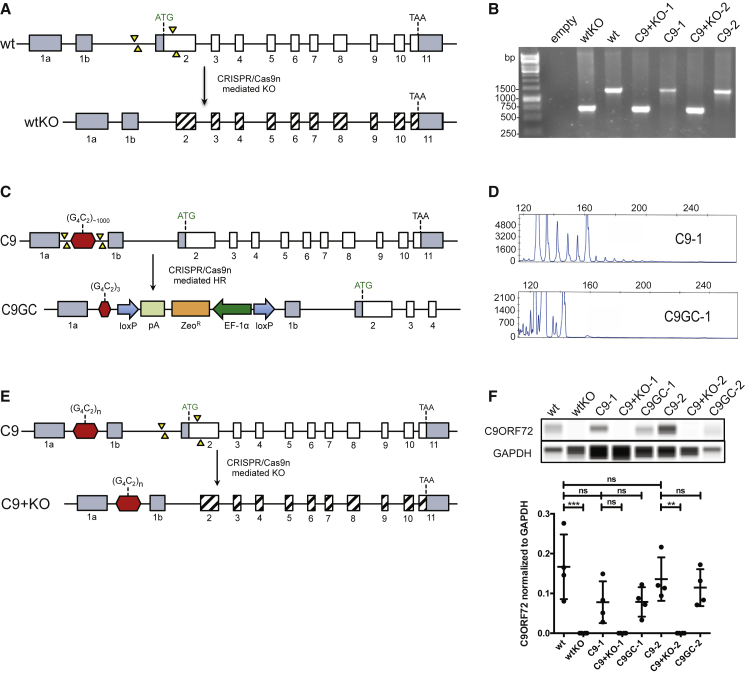
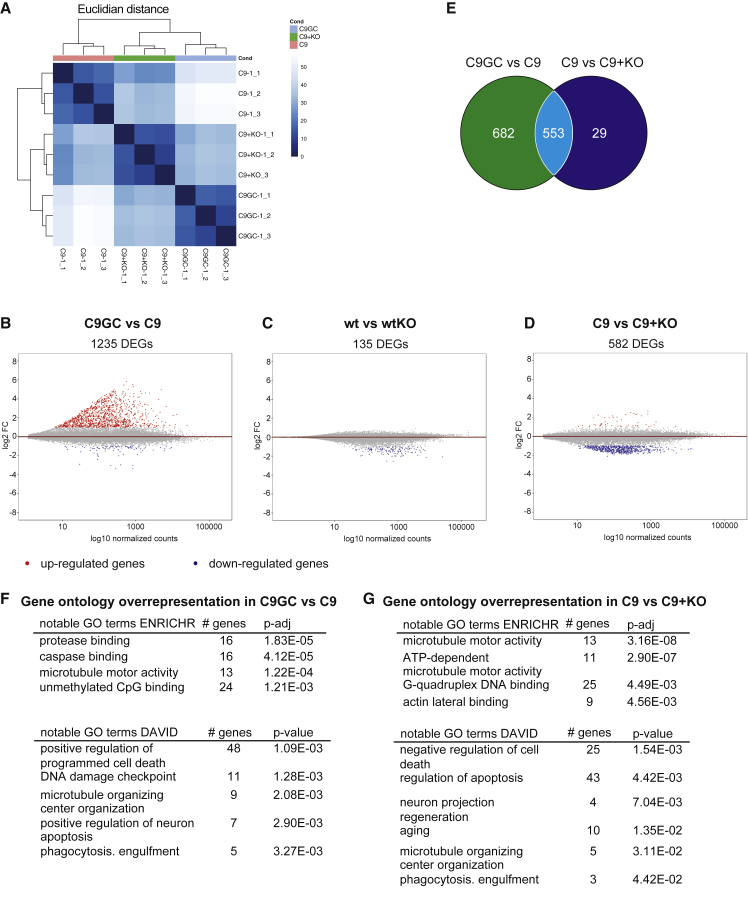
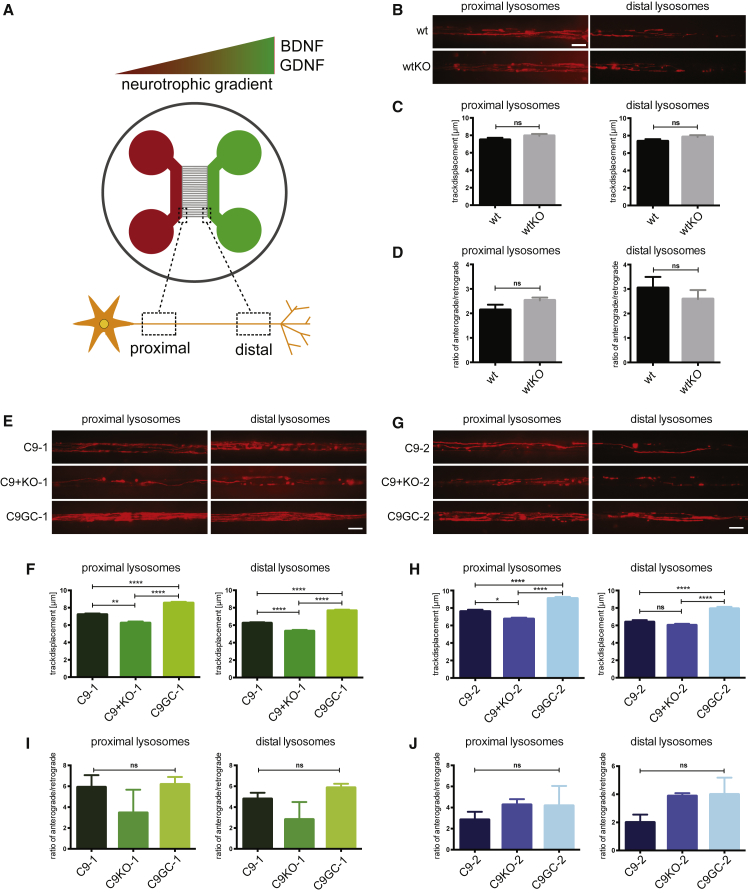
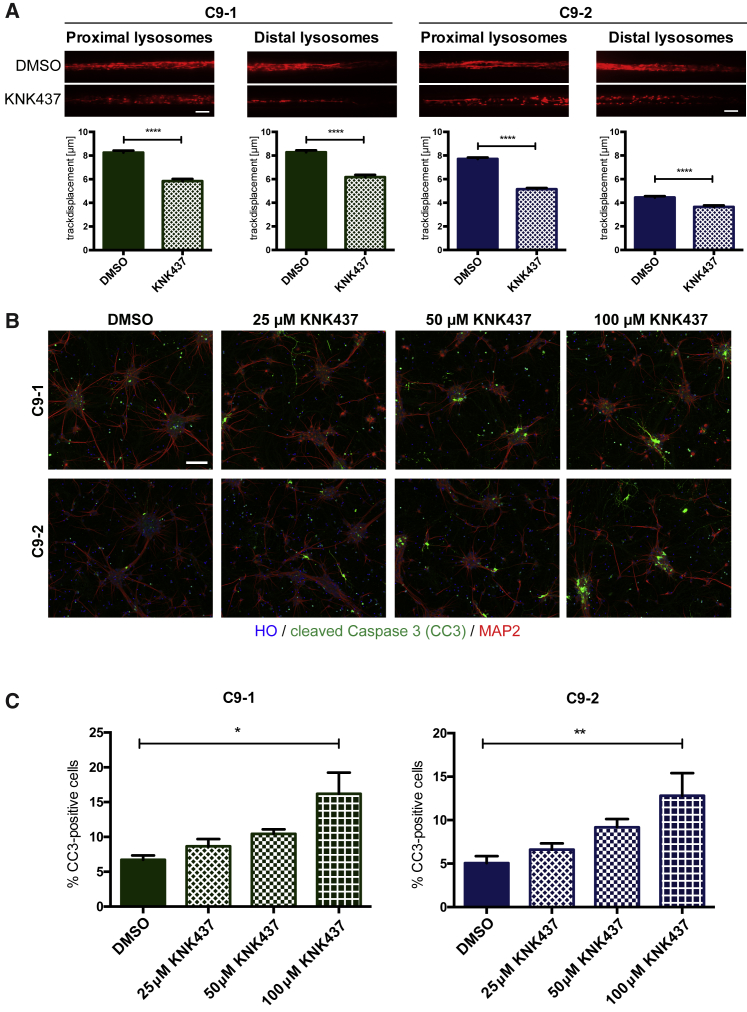
In amyotrophic lateral sclerosis (ALS) motor neurons (MNs) undergo dying-back, where the distal axon degenerates before the soma. The hexanucleotide repeat expansion (HRE) in C9ORF72 is the most common genetic cause of ALS, but the mechanism of pathogenesis is largely unknown with both gain- and loss-of-function mechanisms being proposed. To better understand C9ORF72-ALS pathogenesis, we generated isogenic induced pluripotent stem cells. MNs with HRE in C9ORF72 showed decreased axonal trafficking compared with gene corrected MNs. However, knocking out C9ORF72 did not recapitulate these changes in MNs from healthy controls, suggesting a gain-of-function mechanism. In contrast, knocking out C9ORF72 in MNs with HRE exacerbated axonal trafficking defects and increased apoptosis as well as decreased levels of HSP70 and HSP40, and inhibition of HSPs exacerbated ALS phenotypes in MNs with HRE. Therefore, we propose that the HRE in C9ORF72 induces ALS pathogenesis via a combination of gain- and loss-of-function mechanisms.
Keywords: C9ORF72; HSP40; HSP70; amyotrophic lateral sclerosis; axonal trafficking; disease modeling; gene editing; heat shock proteins; induced pluripotent stem cells.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Ciura S., Lattante S., Le Ber I., Latouche M., Tostivint H., Brice A., Kabashi E. Loss of function of C9orf72 causes motor deficits in a zebrafish model of amyotrophic lateral sclerosis. Ann. Neurol. 2013;74:180–187. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
