

Distinct Mesenchymal Cell Populations Generate the Essential Intestinal BMP Signaling Gradient
- PMID: 32084389
- PMCID: PMC7412576
- DOI: 10.1016/j.stem.2020.01.008
Distinct Mesenchymal Cell Populations Generate the Essential Intestinal BMP Signaling Gradient
Abstract
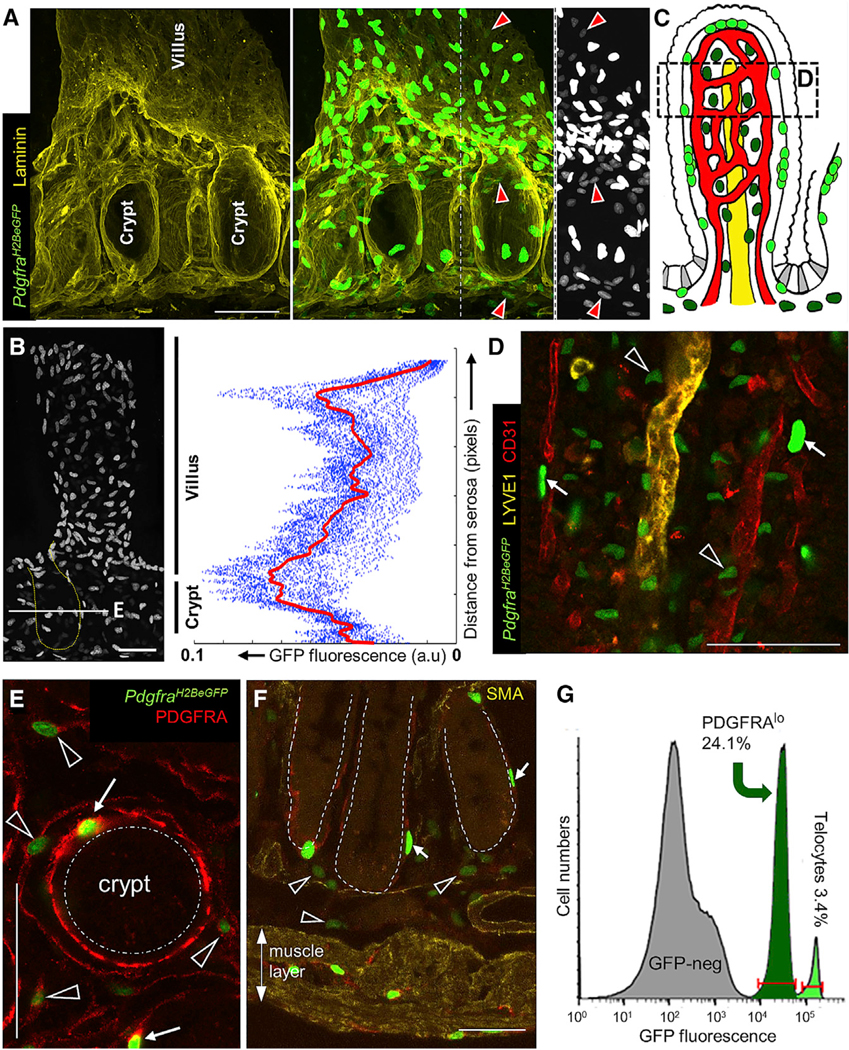
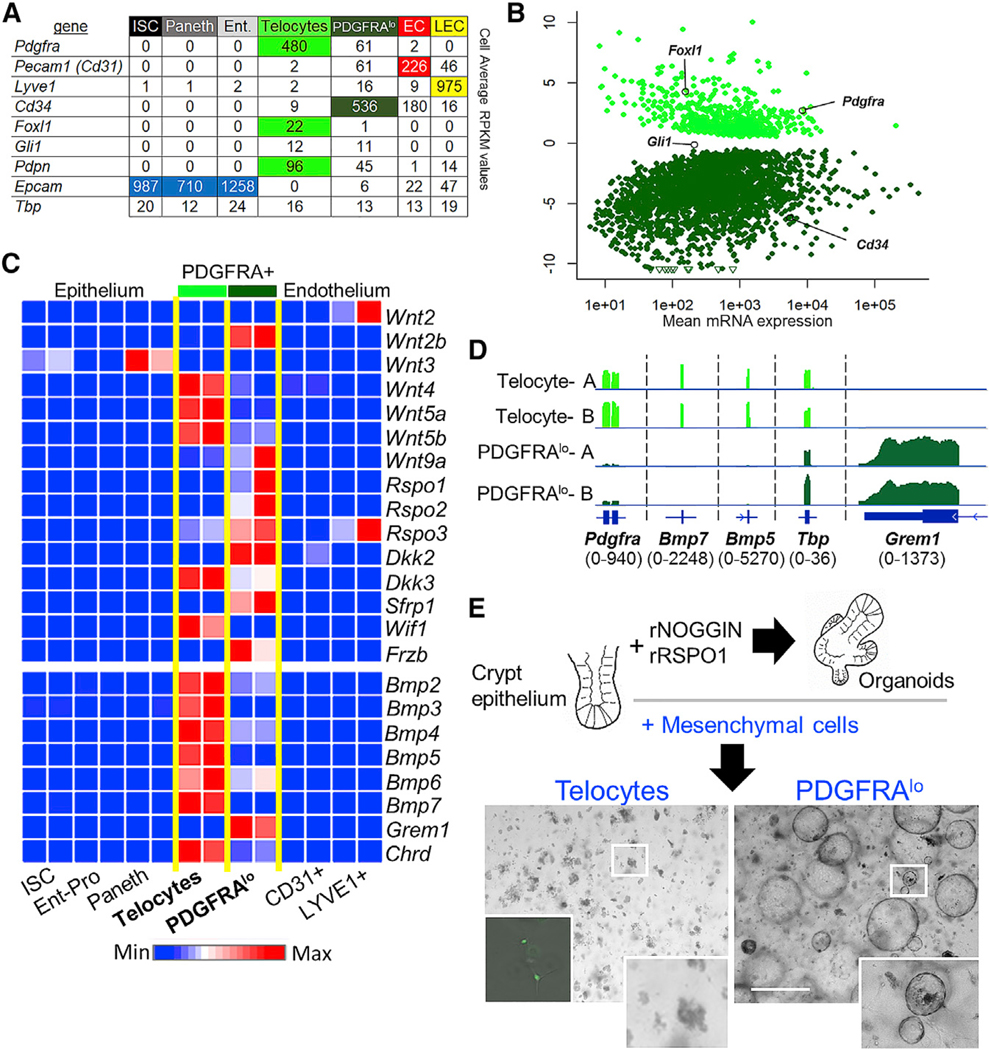
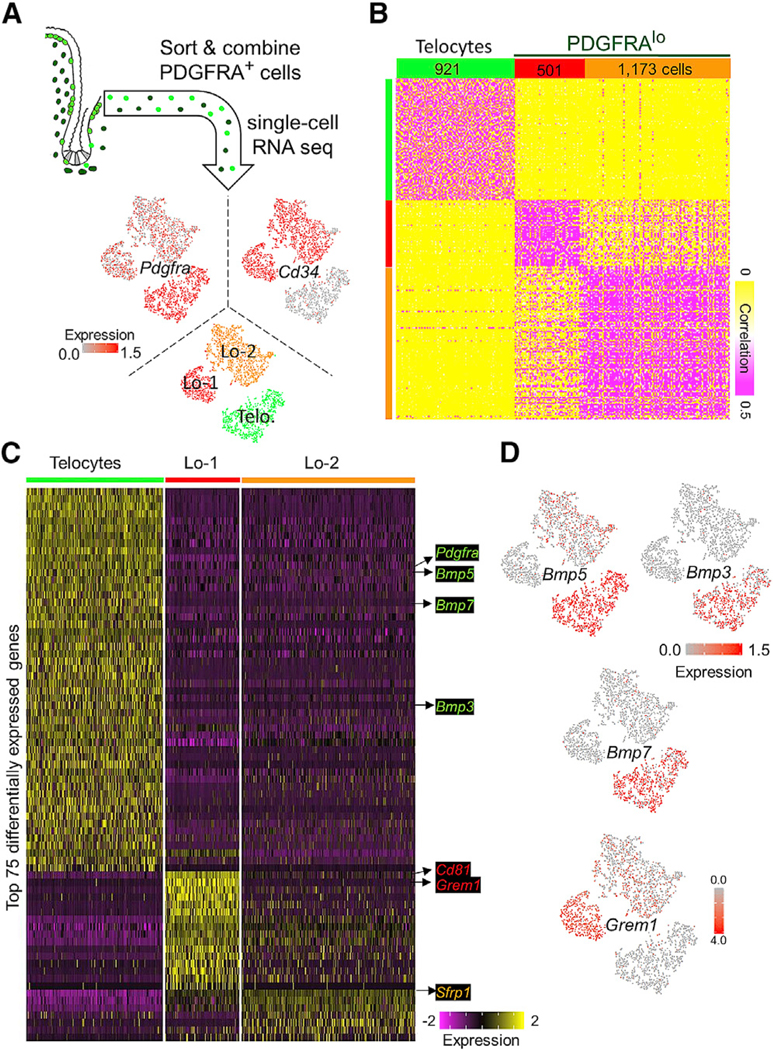
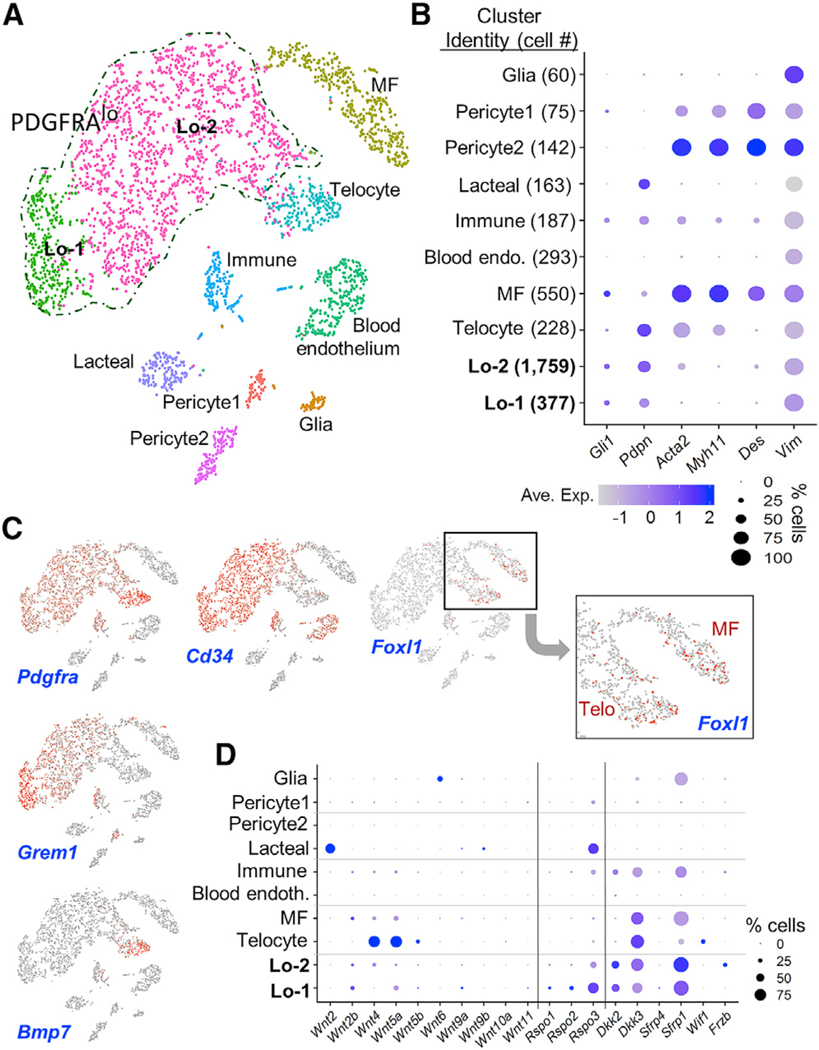
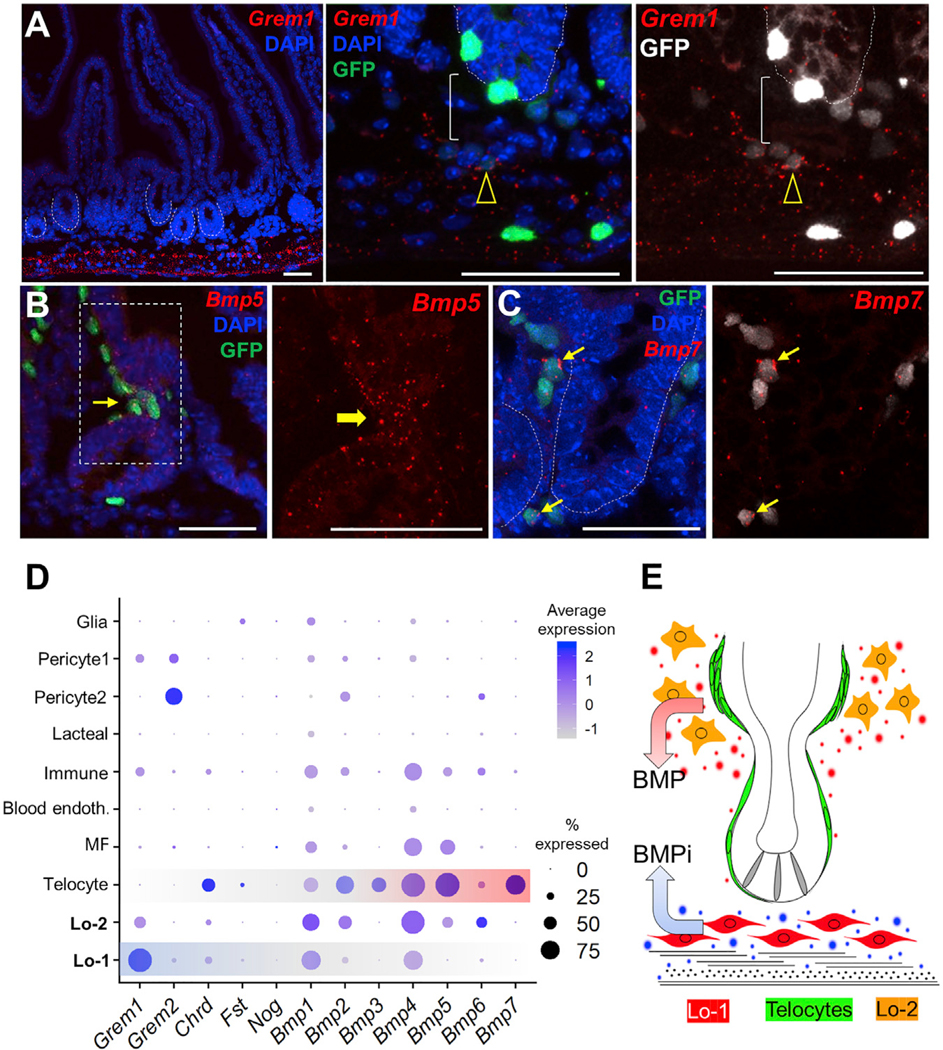
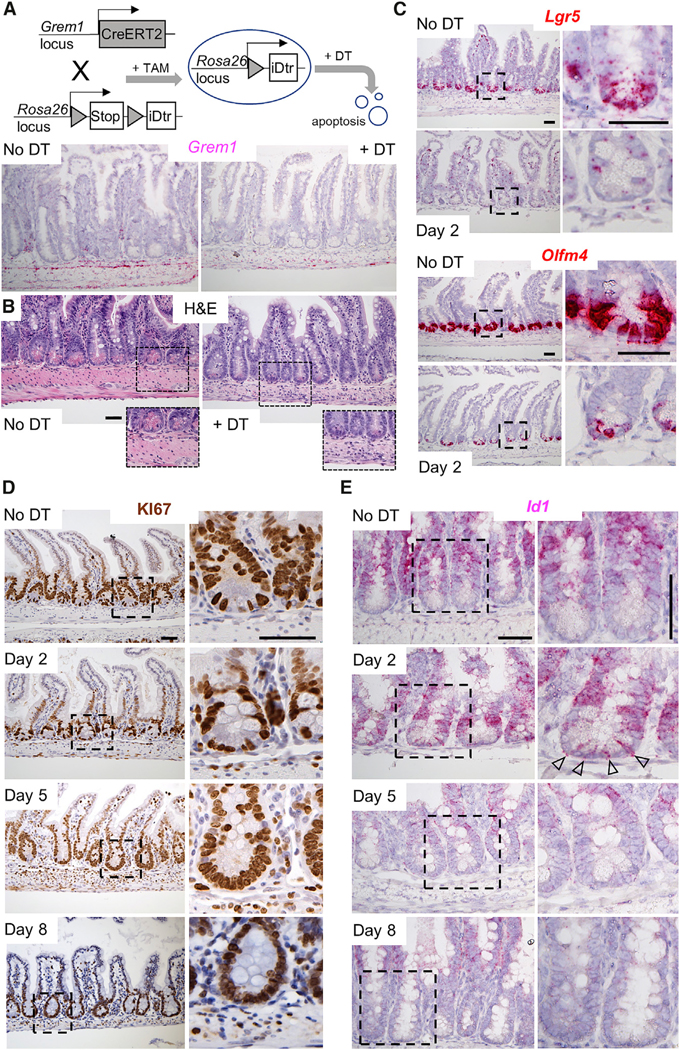
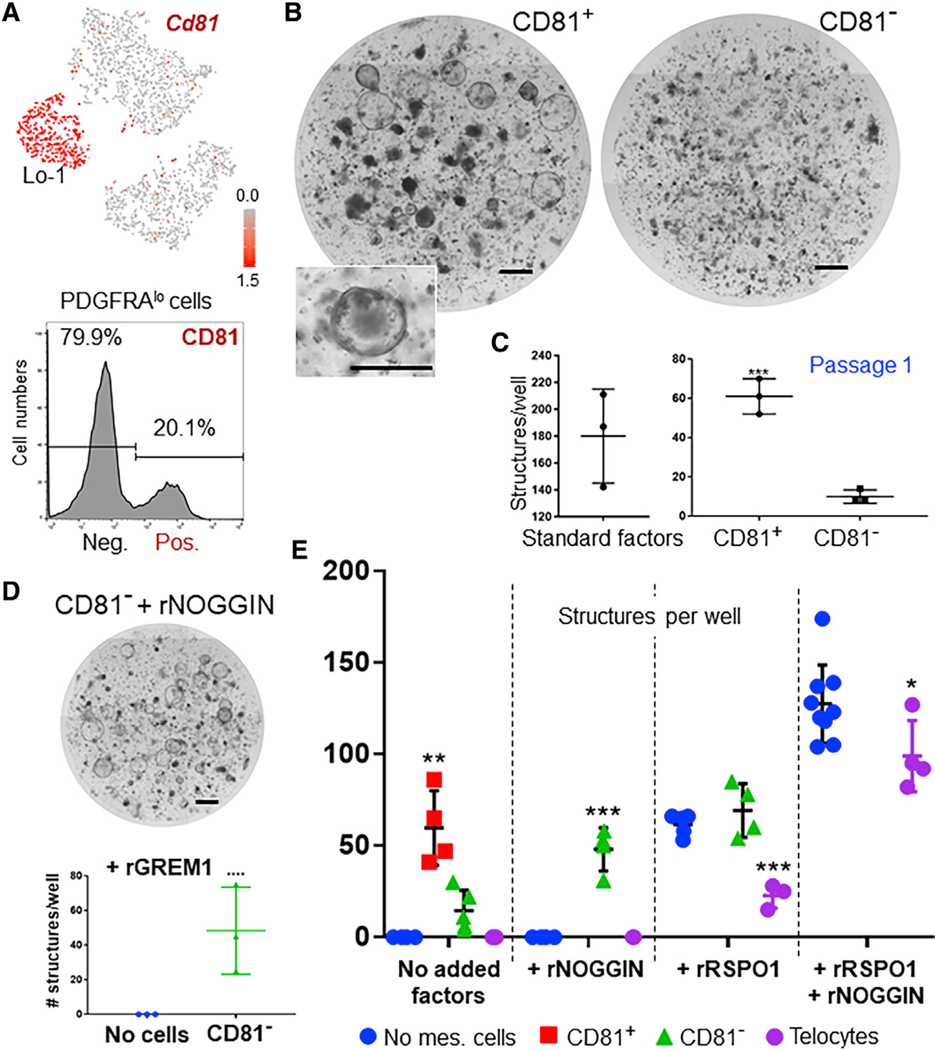
Intestinal stem cells (ISCs) are confined to crypt bottoms and their progeny differentiate near crypt-villus junctions. Wnt and bone morphogenic protein (BMP) gradients drive this polarity, and colorectal cancer fundamentally reflects disruption of this homeostatic signaling. However, sub-epithelial sources of crucial agonists and antagonists that organize this BMP gradient remain obscure. Here, we couple whole-mount high-resolution microscopy with ensemble and single-cell RNA sequencing (RNA-seq) to identify three distinct PDGFRA+ mesenchymal cell types. PDGFRA(hi) telocytes are especially abundant at the villus base and provide a BMP reservoir, and we identified a CD81+ PDGFRA(lo) population present just below crypts that secretes the BMP antagonist Gremlin1. These cells, referred to as trophocytes, are sufficient to expand ISCs in vitro without additional trophic support and contribute to ISC maintenance in vivo. This study reveals intestinal mesenchymal structure at fine anatomic, molecular, and functional detail and the cellular basis for a signaling gradient necessary for tissue self-renewal.
Keywords: epithelium-mesenchyme co-culture; intestinal stem cell niche; mesenchyme; niche signaling gradients; single-cell RNA profiles; telocytes; trophocytes.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests E.E.S., V.N.K., C.C., S.K., F.J.d.S., and S.J.T. are employees of Genentech and own shares in Roche. The other authors declare no competing interests.
Figures
References
-
- Aono A, Hazama M, Notoya K, Taketomi S, Yamasaki H, Tsukuda R, Sasaki S, and Fujisawa Y. (1995). Potent ectopic bone-inducing activity of bone morphogenetic protein-4/7 heterodimer. Biochem. Biophys. Res. Commun 210, 670–677. - PubMed
-
- Batts LE, Polk DB, Dubois RN, and Kulessa H. (2006). Bmp signaling is required for intestinal growth and morphogenesis. Dev. Dyn 235, 1563–1570. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
