

α2δ-2 Protein Controls Structure and Function at the Cerebellar Climbing Fiber Synapse
- PMID: 32086258
- PMCID: PMC7083538
- DOI: 10.1523/JNEUROSCI.1514-19.2020
α2δ-2 Protein Controls Structure and Function at the Cerebellar Climbing Fiber Synapse
Abstract
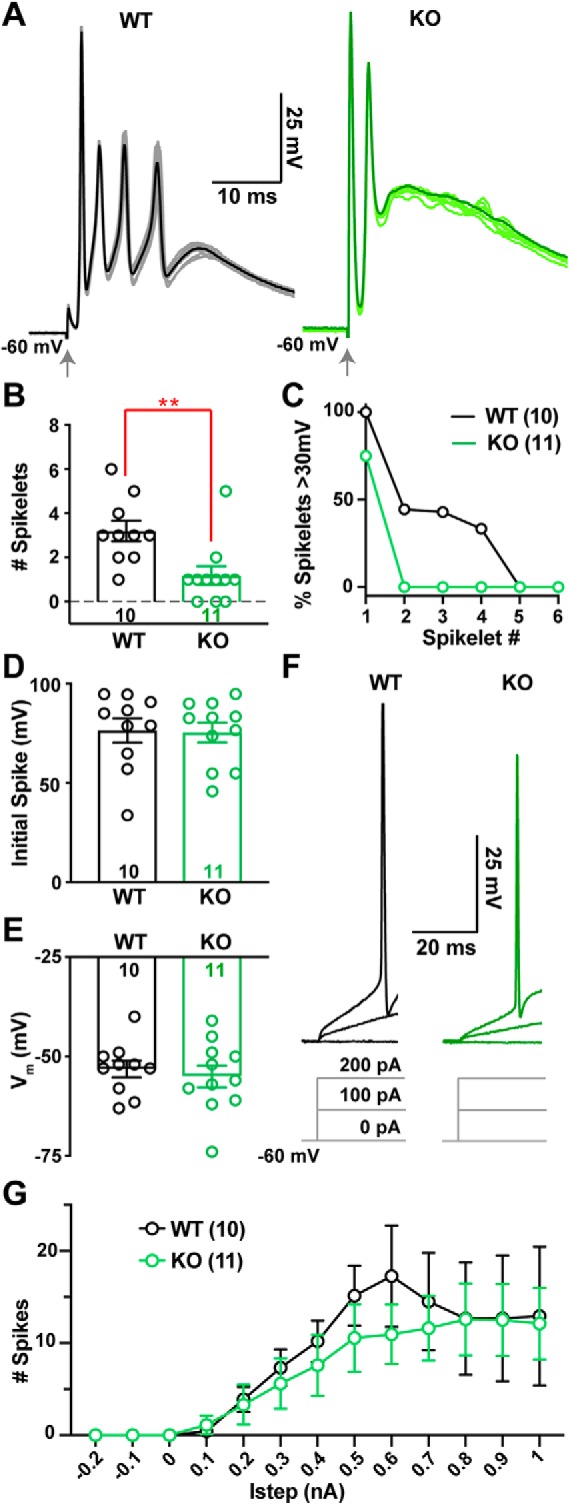
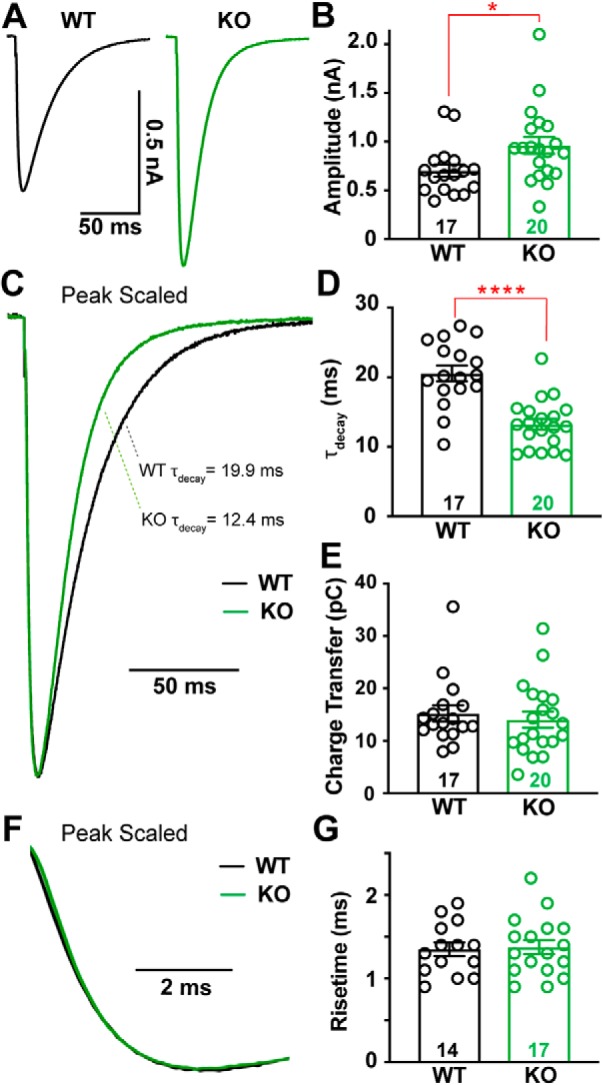
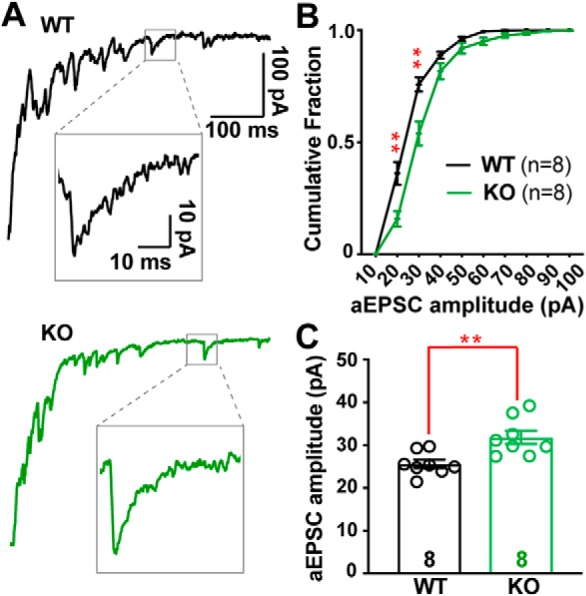
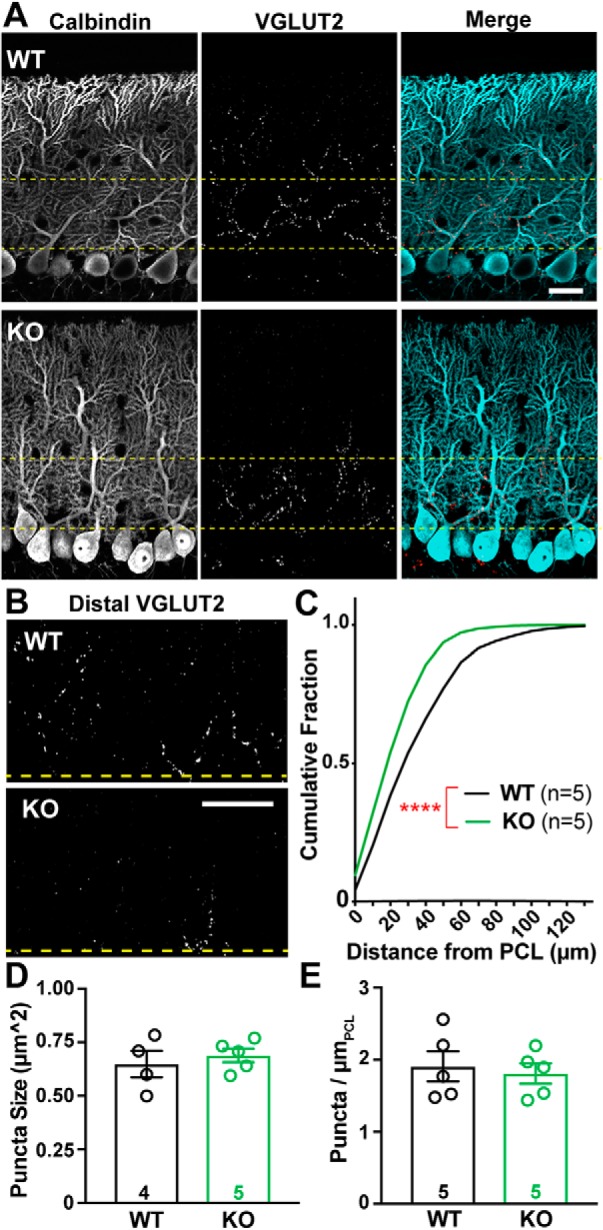
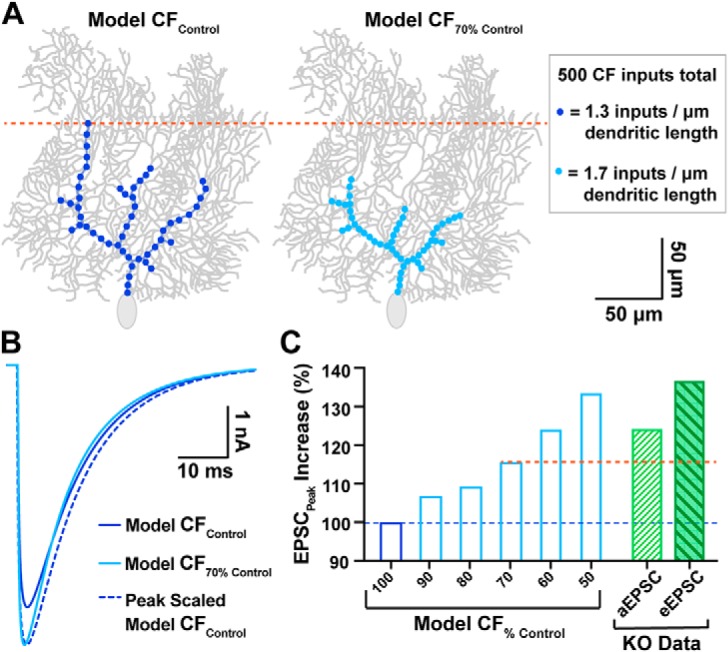
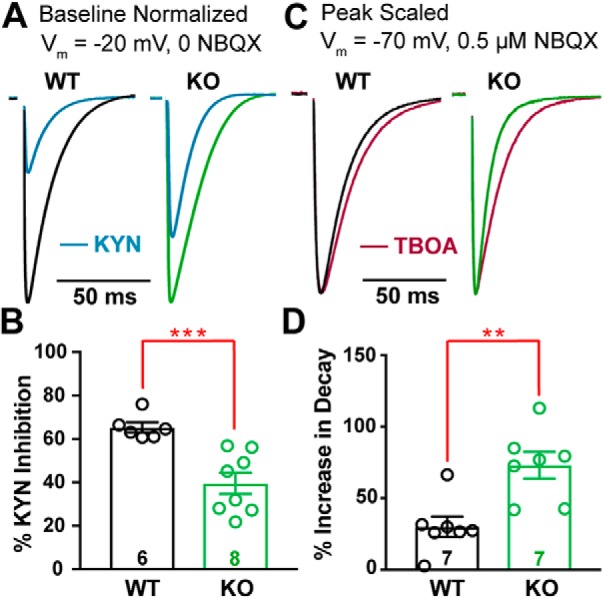
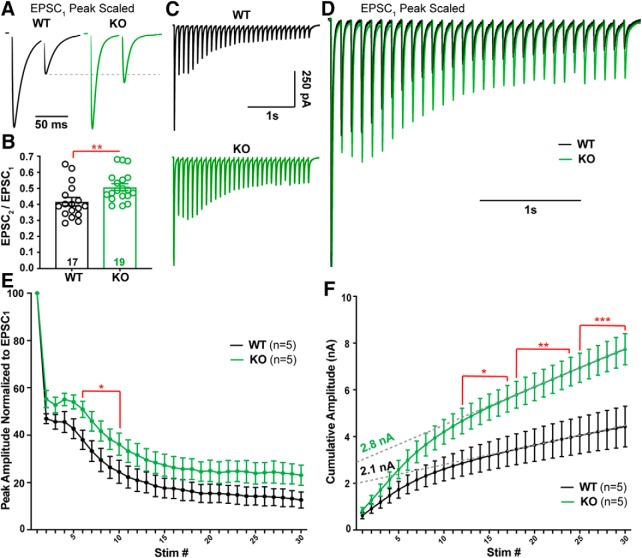
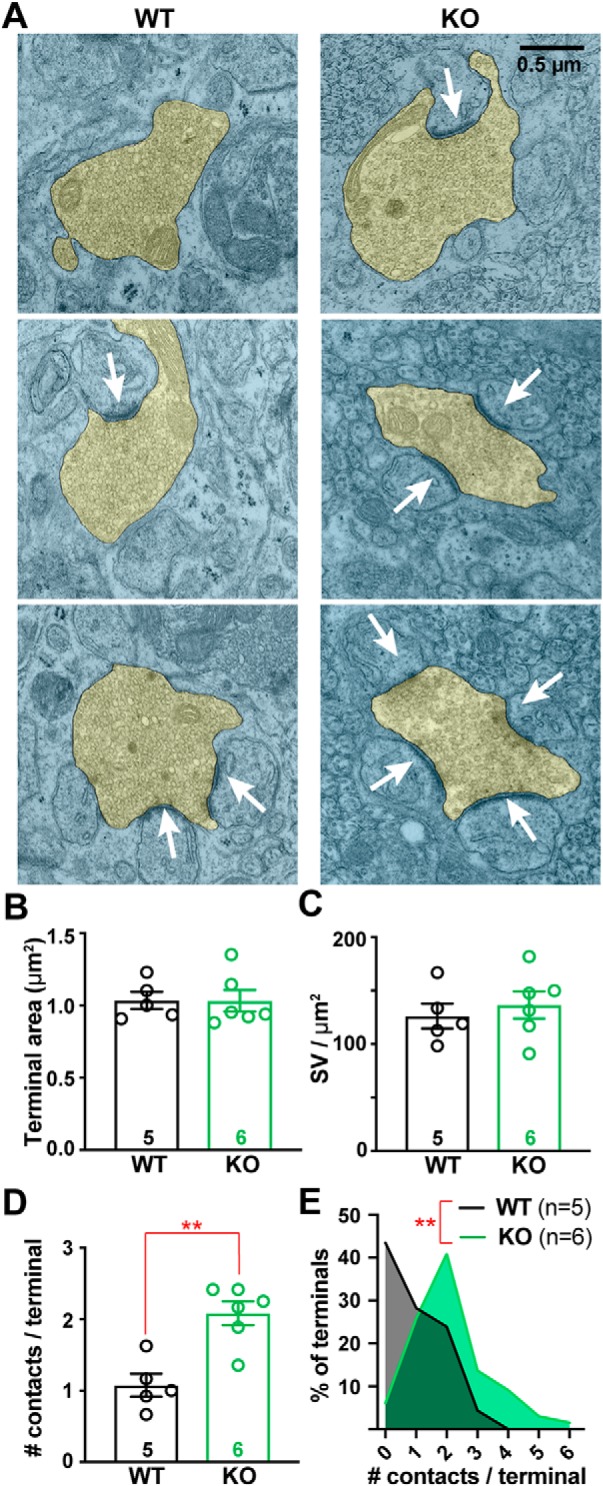
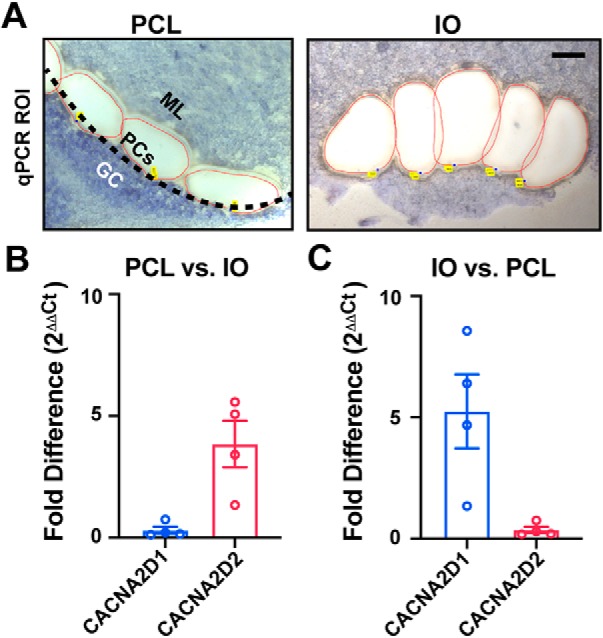
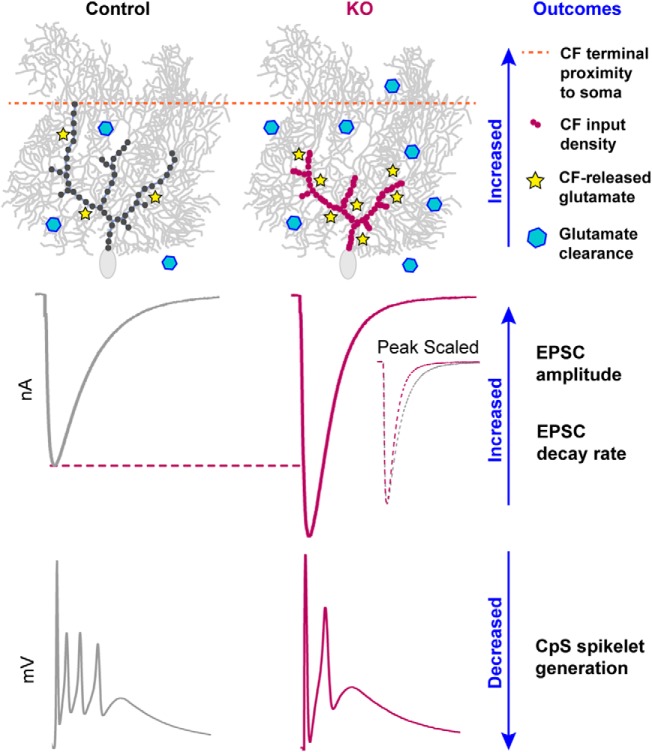
α2δ proteins (Cacna2d1-4) are auxiliary subunits of voltage-dependent calcium channels that also drive synapse formation and maturation. Because cerebellar Purkinje cells (PCs) predominantly, if not exclusively, express one isoform of this family, α2δ-2 (Cacna2d2), we used PCs as a model system to examine roles of α2δ in excitatory synaptic function in male and female Cacna2d2 knock-out (KO) mice. Whole-cell recordings of PCs from acute cerebellar slices revealed altered climbing fiber (CF)-evoked complex spike generation, as well as increased amplitude and faster decay of CF-evoked EPSCs. CF terminals in the KO were localized more proximally on PC dendrites, as indicated by VGLUT2+ immunoreactive puncta, and computational modeling demonstrated that the increased EPSC amplitude can be partly attributed to the more proximal location of CF terminals. In addition, CFs in KO mice exhibited increased multivesicular transmission, corresponding to greater sustained responses during repetitive stimulation, despite a reduction in the measured probability of release. Electron microscopy demonstrated that mutant CF terminals had twice as many vesicle release sites, providing a morphologic explanation for the enhanced glutamate release. Though KO CFs evoked larger amplitude EPSCs, the charge transfer was the same as wild-type as a result of increased glutamate reuptake, producing faster decay kinetics. Together, the larger, faster EPSCs in the KO explain the altered complex spike responses, which degrade information transfer from PCs and likely contribute to ataxia in Cacna2d2 KO mice. Our results also illustrate the multidimensional synaptic roles of α2δ proteins.SIGNIFICANCE STATEMENT α2δ proteins (Cacna2d1-4) regulate synaptic transmission and synaptogenesis, but coexpression of multiple α2δ isoforms has obscured a clear understanding of how various α2δ proteins control synaptic function. We focused on roles of the α2δ-2 protein (Cacna2d2), the deletion of which causes cerebellar ataxia and epilepsy in mice and humans. Because cerebellar Purkinje cells (PCs) only express this single isoform, we studied excitatory climbing fiber synaptic function onto PCs in Cacna2d2 KO mice. Using optical and electrophysiological analysis, we provide a detailed description of the changes in PCs lacking α2δ-2, and provide a comprehensive mechanistic explanation for how functional synaptic phenotypes contribute to the altered cerebellar output.
Keywords: CACNA2D2; Purkinje cell; alpha2delta proteins; calcium channel; climbing fiber.
Copyright © 2020 the authors.
Figures
References
-
- Barclay J, Balaguero N, Mione M, Ackerman SL, Letts VA, Brodbeck J, Canti C, Meir A, Page KM, Kusumi K, Perez-Reyes E, Lander ES, Frankel WN, Gardiner RM, Dolphin AC, Rees M (2001) Ducky mouse phenotype of epilepsy and ataxia is associated with mutations in the Cacna2d2 gene and decreased calcium channel current in cerebellar Purkinje cells. J Neurosci 21:6095–6104. 10.1523/JNEUROSCI.21-16-06095.2001 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials