

Pathogen-Associated Molecular Pattern-Triggered Immunity Involves Proteolytic Degradation of Core Nonsense-Mediated mRNA Decay Factors During the Early Defense Response
- PMID: 32086363
- PMCID: PMC7145493
- DOI: 10.1105/tpc.19.00631
Pathogen-Associated Molecular Pattern-Triggered Immunity Involves Proteolytic Degradation of Core Nonsense-Mediated mRNA Decay Factors During the Early Defense Response
Abstract
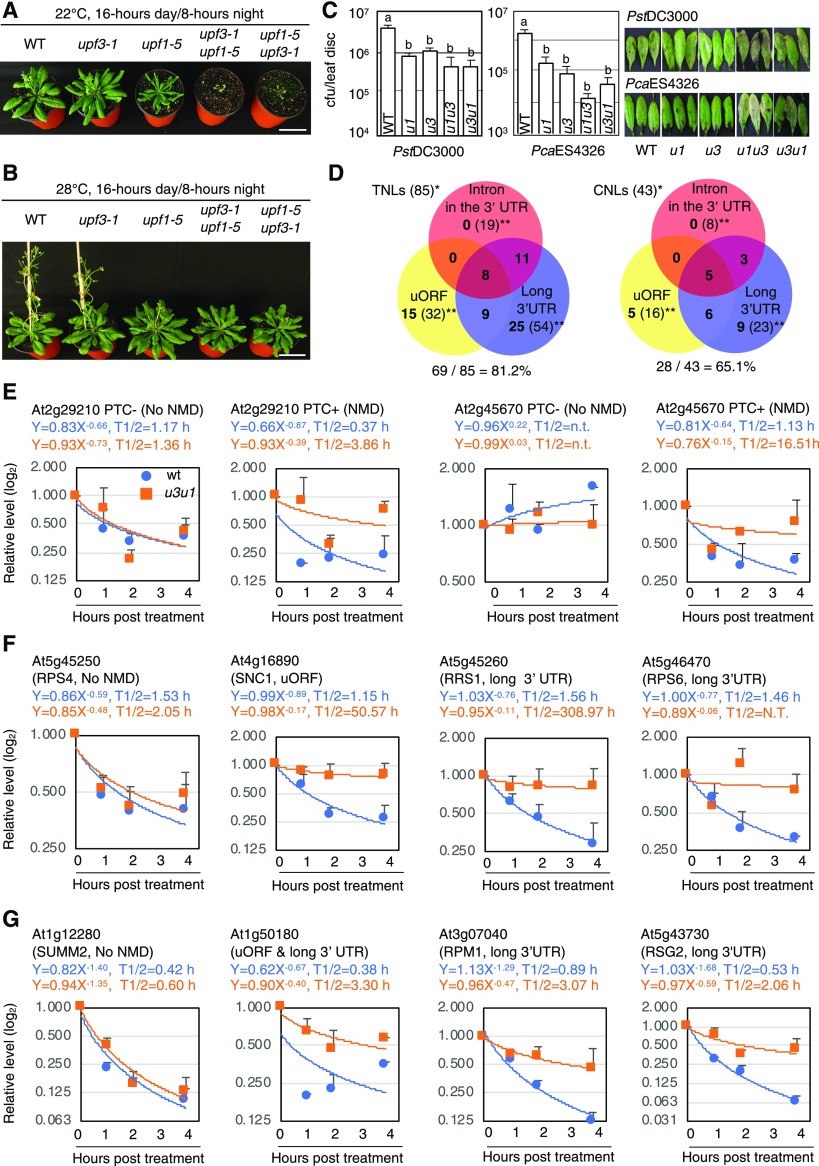
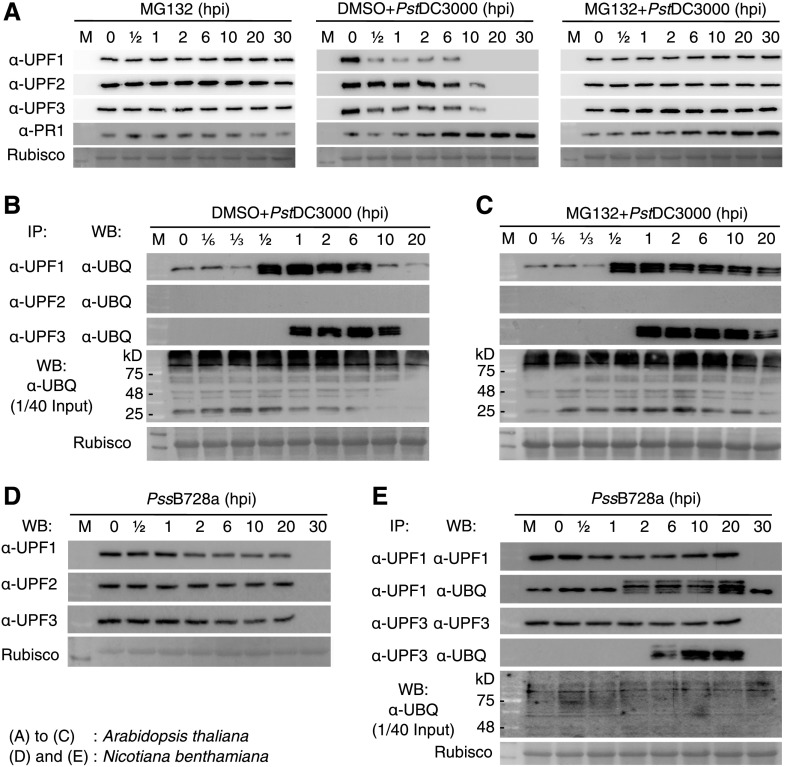
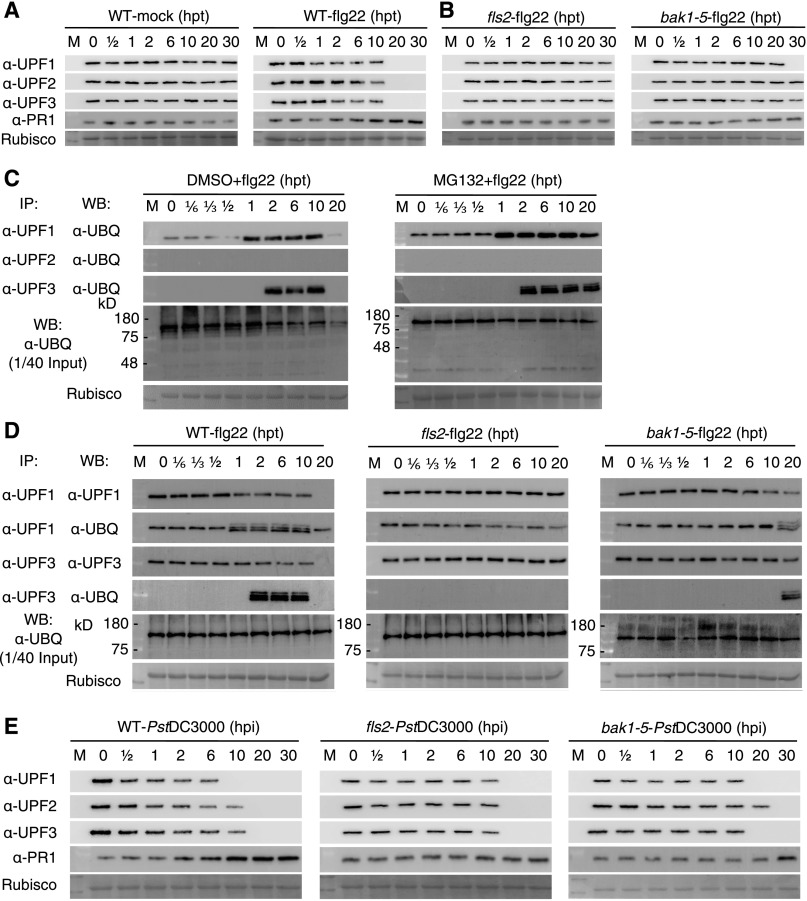
Nonsense-mediated mRNA decay (NMD), an mRNA quality control process, is thought to function in plant immunity. A subset of fully spliced (FS) transcripts of Arabidopsis (Arabidopsis thaliana) resistance (R) genes are upregulated during bacterial infection. Here, we report that 81.2% and 65.1% of FS natural TIR-NBS-LRR (TNL) and CC-NBS-LRR transcripts, respectively, retain characteristics of NMD regulation, as their transcript levels could be controlled posttranscriptionally. Both bacterial infection and the perception of bacteria by pattern recognition receptors initiated the destruction of core NMD factors UP-FRAMESHIFT1 (UPF1), UPF2, and UPF3 in Arabidopsis within 30 min of inoculation via the independent ubiquitination of UPF1 and UPF3 and their degradation via the 26S proteasome pathway. The induction of UPF1 and UPF3 ubiquitination was delayed in mitogen-activated protein kinase3 (mpk3) and mpk6, but not in salicylic acid-signaling mutants, during the early immune response. Finally, previously uncharacterized TNL-type R transcripts accumulated in upf mutants and conferred disease resistance to infection with a virulent Pseudomonas strain in plants. Our findings demonstrate that NMD is one of the main regulatory processes through which PRRs fine-tune R transcript levels to reduce fitness costs and achieve effective immunity.
© 2020 American Society of Plant Biologists. All rights reserved.
Figures





Comment in
-
Security Notice: This Plant Immunity Is under mRNA Surveillance.Plant Cell. 2020 Apr;32(4):803-804. doi: 10.1105/tpc.20.00169. Epub 2020 Feb 28. Plant Cell. 2020. PMID: 32111665 Free PMC article. No abstract available.
References
-
- Alcázar R., Parker J.E.(2011). The impact of temperature on balancing immune responsiveness and growth in Arabidopsis. Trends Plant Sci. 16: 666–675. - PubMed
-
- Arciga-Reyes L., Wootton L., Kieffer M., Davies B.(2006). UPF1 is required for nonsense-mediated mRNA decay (NMD) and RNAi in Arabidopsis. Plant J. 47: 480–489. - PubMed
-
- Boccara M., Sarazin A., Thiébeauld O., Jay F., Voinnet O., Navarro L., Colot V.(2015). Correction: The Arabidopsis miR472-RDR6 silencing pathway modulates PAMP- and effector-triggered immunity through the post-transcriptional control of disease resistance genes. PLoS Pathog. 11: e1004814. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
