

Sustained hyperammonemia induces TNF-a IN Purkinje neurons by activating the TNFR1-NF-κB pathway
- PMID: 32087723
- PMCID: PMC7035786
- DOI: 10.1186/s12974-020-01746-z
Sustained hyperammonemia induces TNF-a IN Purkinje neurons by activating the TNFR1-NF-κB pathway
Abstract
Background: Patients with liver cirrhosis may develop hepatic encephalopathy. Rats with chronic hyperammonemia exhibit neurological alterations mediated by peripheral inflammation and neuroinflammation. Motor incoordination is due to increased TNF-a levels and activation of its receptor TNFR1 in the cerebellum. The aims were to assess (a) whether peripheral inflammation is responsible for TNF-a induction in hyperammonemic rats, (b) the cell type(s) in which TNF-a is increased, (c) whether this increase is associated with increased nuclear NF-κB and TNFR1 activation, (d) the time course of TNF-a induction, and (e) if TNF-a is induced in the Purkinje neurons of patients who die with liver cirrhosis.
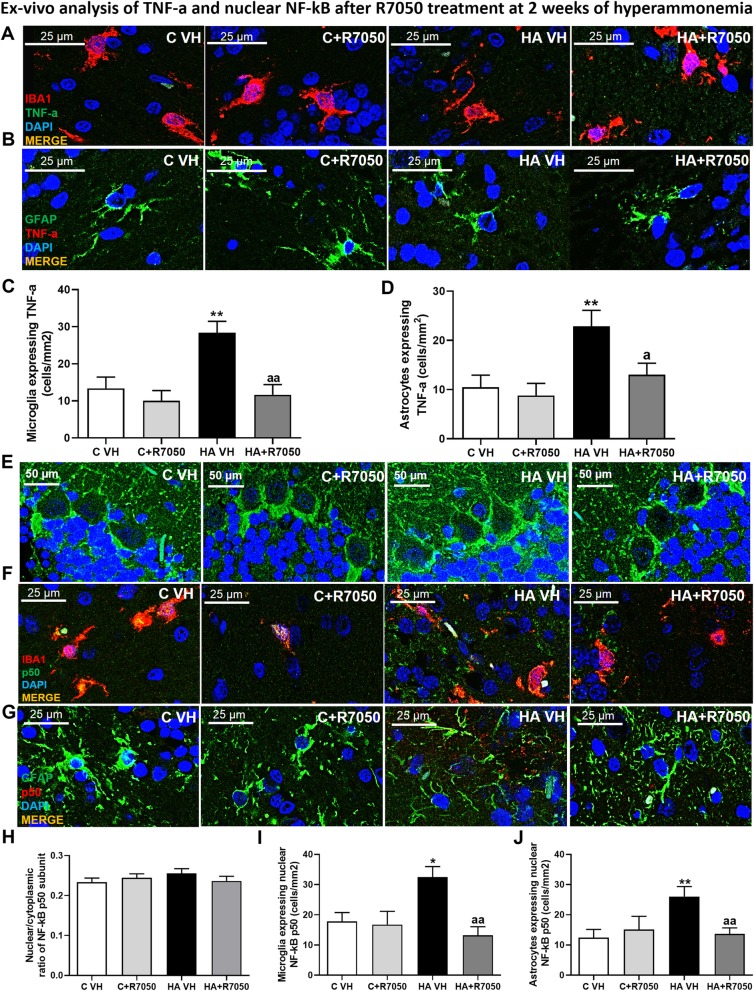
Methods: We analyzed the level of TNF-a mRNA and NF-κB in microglia, astrocytes, and Purkinje neurons in the cerebellum after 1, 2, and 4 weeks of hyperammonemia. We assessed whether preventing peripheral inflammation by administering an anti-TNF-a antibody prevents TNF-a induction. We tested whether TNF-a induction is reversed by R7050, which inhibits the TNFR1-NF-κB pathway, in ex vivo cerebellar slices.
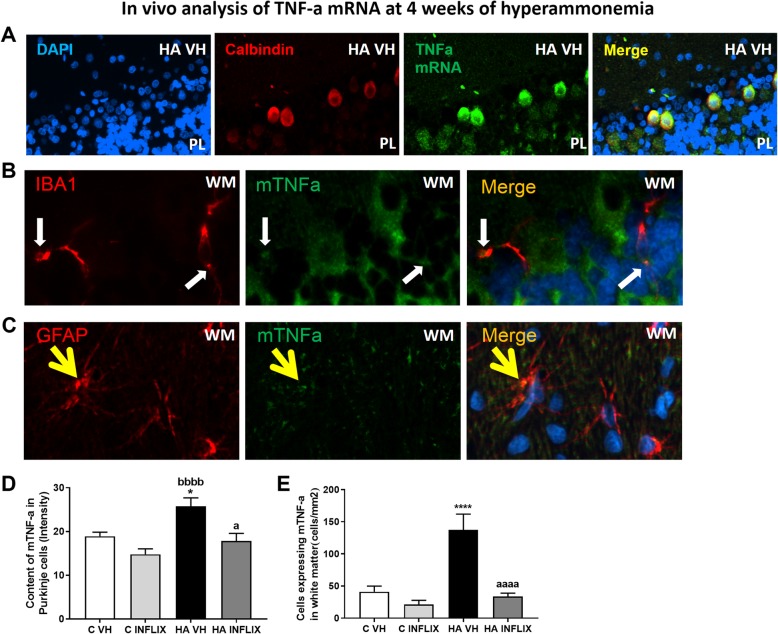
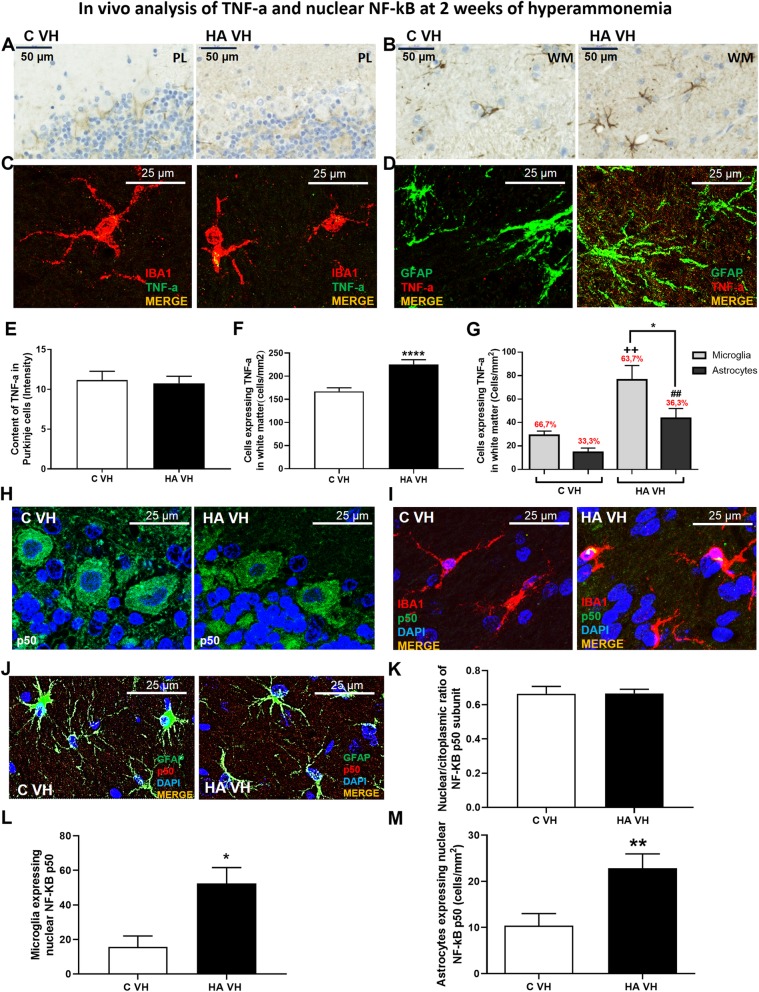
Results: Hyperammonemia induced microglial and astrocyte activation at 1 week. This was followed by TNF-a induction in both glial cell types at 2 weeks and in Purkinje neurons at 4 weeks. The level of TNF-a mRNA increased in parallel with the TNF-a protein level, indicating that TNF-a was synthesized in Purkinje cells. This increase was associated with increased NF-κB nuclear translocation. The nuclear translocation of NF-κB and the increase in TNF-a were reversed by R7050, indicating that they were mediated by the activation of TNFR1. Preventing peripheral inflammation with an anti-TNF-a antibody prevents TNF-a induction.
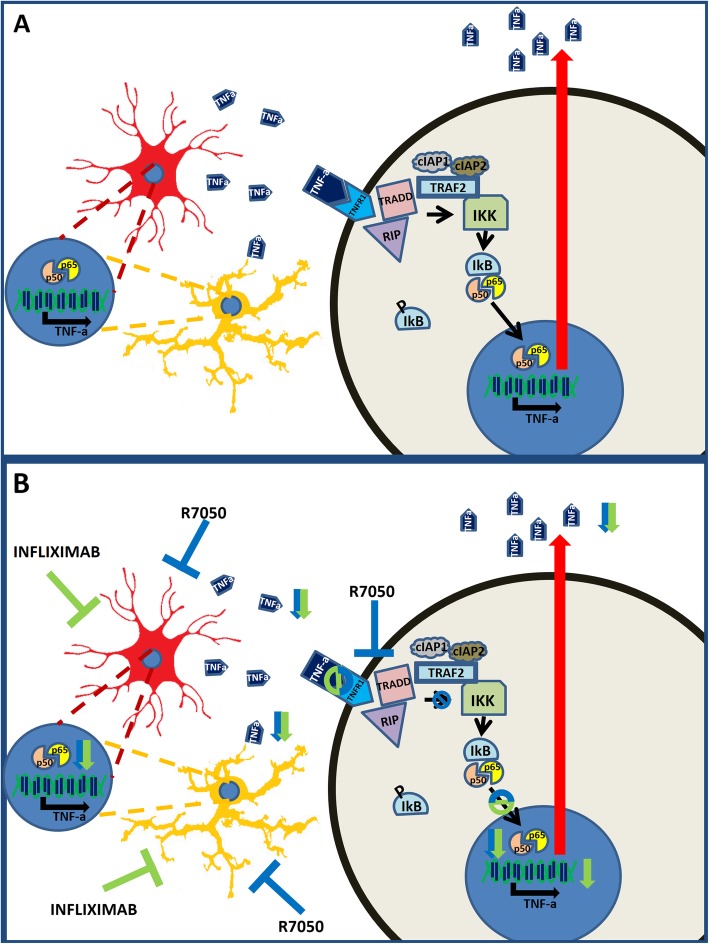
Conclusion: Sustained (4 weeks) but not short-term hyperammonemia induces TNF-a in Purkinje neurons in rats. This is mediated by peripheral inflammation. TNF-a is also increased in the Purkinje neurons of patients who die with liver cirrhosis. The results suggest that hyperammonemia induces TNF-a in glial cells and that TNF-a released by glial cells activates TNFR1 in Purkinje neurons, leading to NF-κB nuclear translocation and the induction of TNF-a expression, which may contribute to the neurological alterations observed in hyperammonemia and hepatic encephalopathy.
Keywords: Hyperammonemia; Neuroinflammation; Purkinje neurons; TNF-a; TNFR1.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures








References
-
- Cabrera-Pastor A, Arenas YM, Taoro-Gonzalez L, Montoliu C, Felipo V. Chronic hyperammonemia alters extracellular glutamate, glutamine and GABA and membrane expression of their transporters in rat cerebellum. Modulation by extracellular cGMP. Neuropharmacology. 2019; - PubMed
MeSH terms
Substances
Grants and funding
- SAF2014-51851-R and SAF2017-82917-R/Ministerio de Ciencia e Innovación
- PROMETEOII/2014/033 and PROMETEOII/2018/051/Conselleria d'Educació, Investigació, Cultura i Esport
- PROMETEOII/2014/033 and PROMETEOII/2018/051 SAF2014-51851-R and SAF2017-82917-R/European Regional Development Funds
- PRE2018-084770/Ministerio de Ciencia e Innovación, Spain
- APOSTD/2018/002/Generalitat Valenciana
LinkOut - more resources
Full Text Sources
