

Insights into the Structure, Function, and Dynamics of the Bacterial Cytokinetic FtsZ-Ring
- PMID: 32092282
- PMCID: PMC8610174
- DOI: 10.1146/annurev-biophys-121219-081703
Insights into the Structure, Function, and Dynamics of the Bacterial Cytokinetic FtsZ-Ring
Abstract
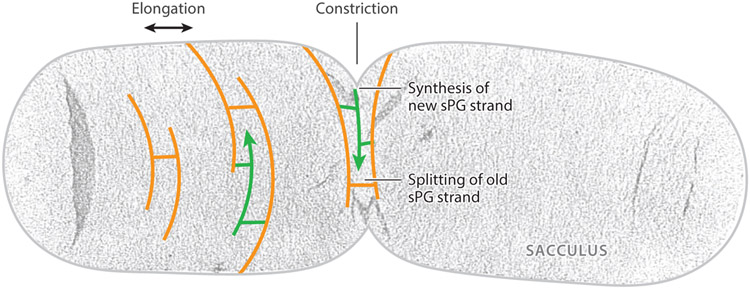
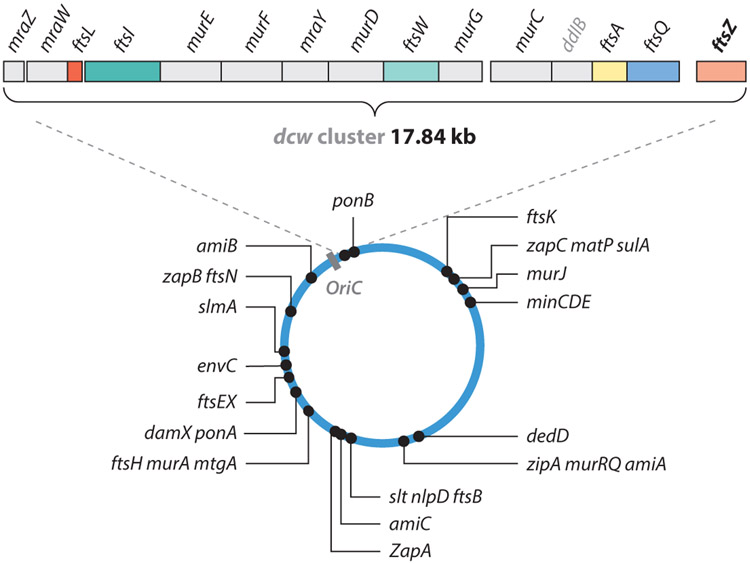
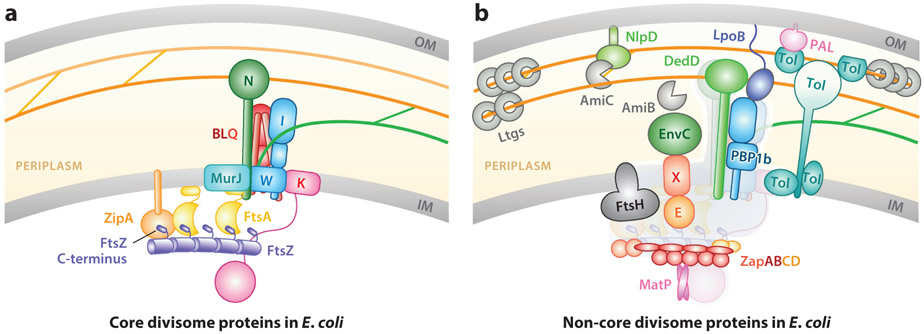
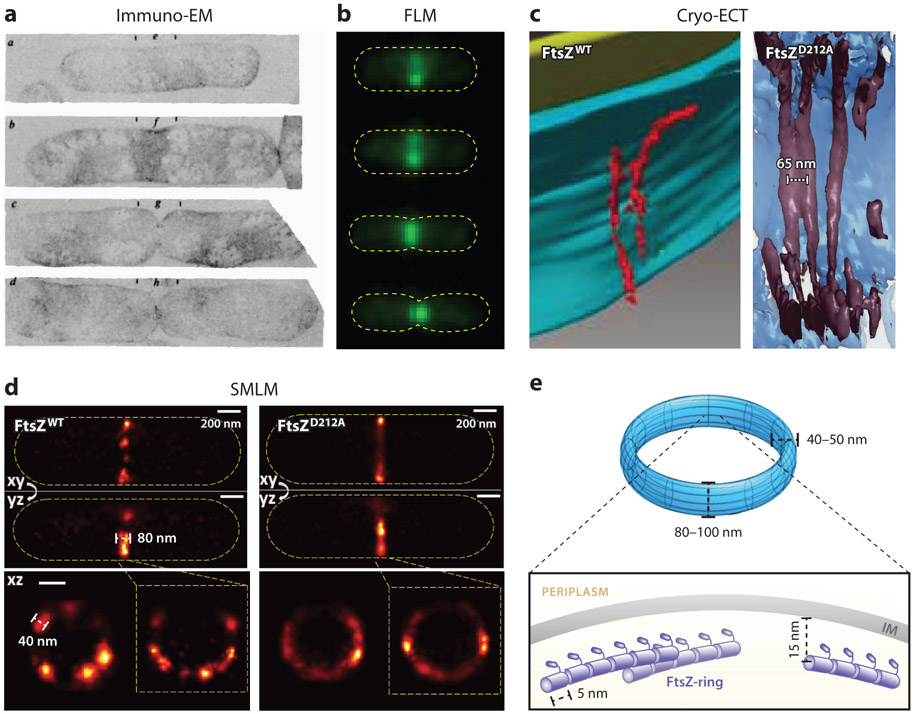
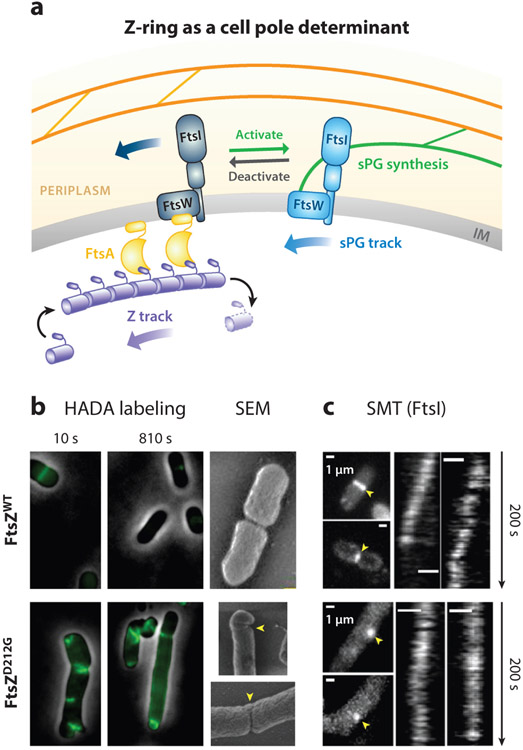
The FtsZ protein is a highly conserved bacterial tubulin homolog. In vivo, the functional form of FtsZ is the polymeric, ring-like structure (Z-ring) assembled at the future division site during cell division. While it is clear that the Z-ring plays an essential role in orchestrating cytokinesis, precisely what its functions are and how these functions are achieved remain elusive. In this article, we review what we have learned during the past decade about the Z-ring's structure, function, and dynamics, with a particular focus on insights generated by recent high-resolution imaging and single-molecule analyses. We suggest that the major function of the Z-ring is to govern nascent cell pole morphogenesis by directing the spatiotemporal distribution of septal cell wall remodeling enzymes through the Z-ring's GTP hydrolysis-dependent treadmilling dynamics. In this role, FtsZ functions in cell division as the counterpart of the cell shape-determining actin homolog MreB in cell elongation.
Keywords: FtsZ; bacterial cell division; cell wall constriction; cytoskeleton dynamics; septum synthesis; single-molecule imaging.
Figures
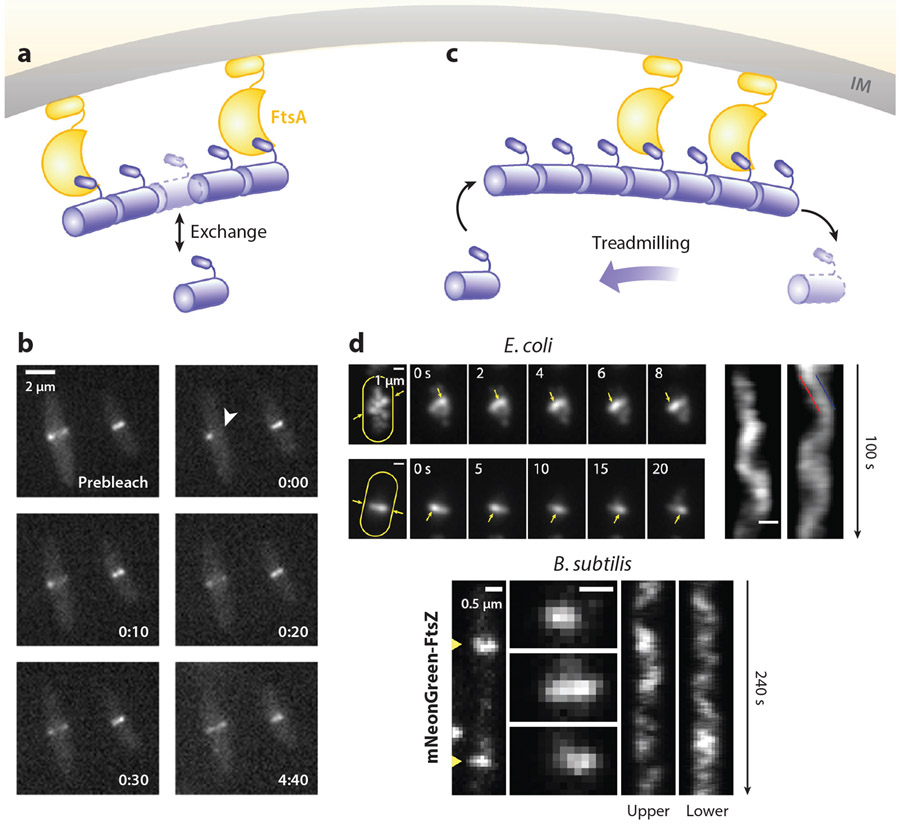
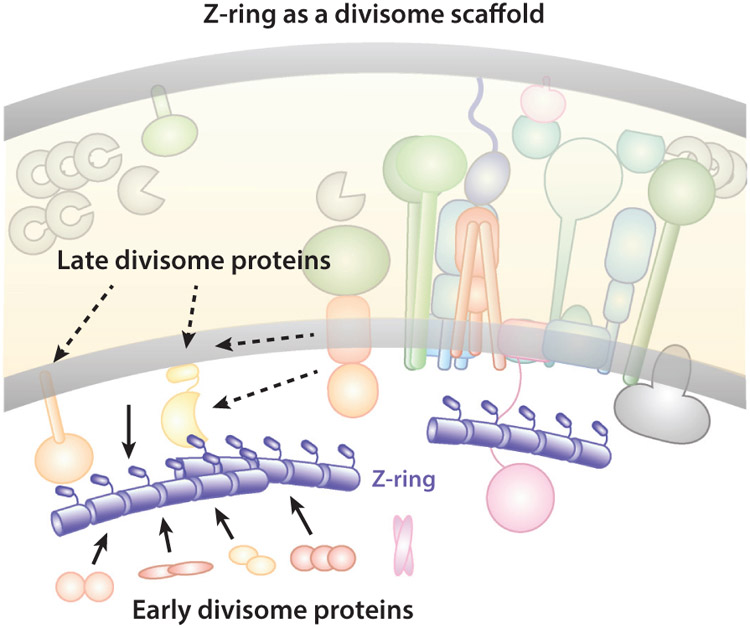
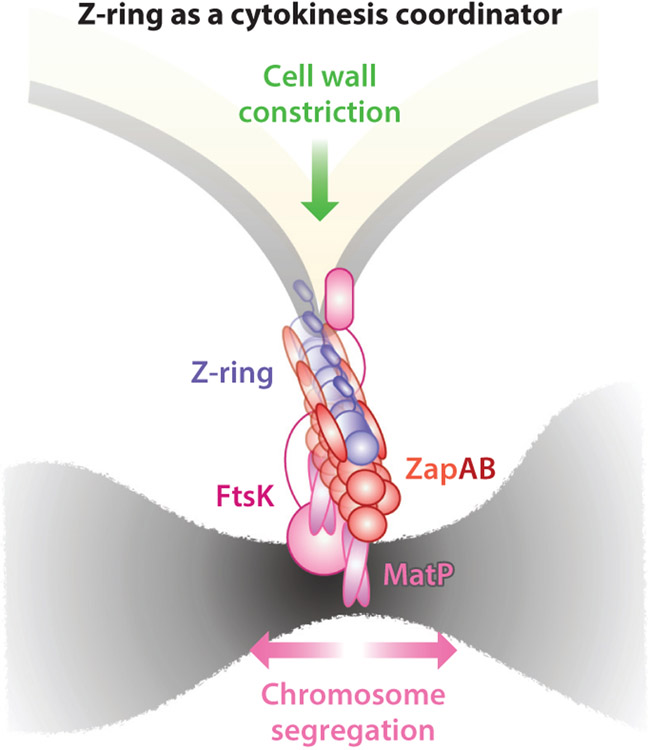
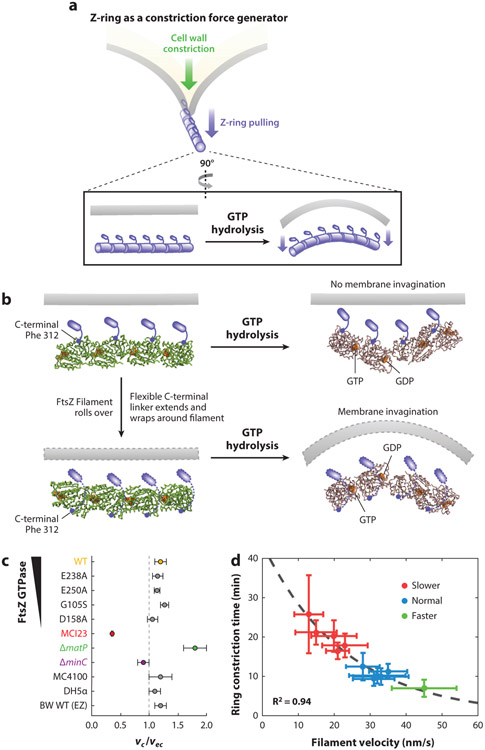
References
-
- Aaron M, Charbon G, Lam H, Schwarz H, Vollmer W, Jacobs-Wagner C. 2007. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol. Microbiol 64:938–52 - PubMed
-
- Aarsman ME, Piette A, Fraipont C, Vinkenvleugel TM, Nguyen-Distèche M, den Blaauwen T. 2005. Maturation of the Escherichia coli divisome occurs in two steps. Mol. Microbiol 55:1631–45 - PubMed
-
- Adams DW, Errington J. 2009. Bacterial cell division: assembly, maintenance and disassembly of the Z ring. Nat. Rev. Microbiol 7:642–53 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
