

NADPH Oxidase 2 Mediates Myocardial Oxygen Wasting in Obesity
- PMID: 32093119
- PMCID: PMC7070669
- DOI: 10.3390/antiox9020171
NADPH Oxidase 2 Mediates Myocardial Oxygen Wasting in Obesity
Abstract
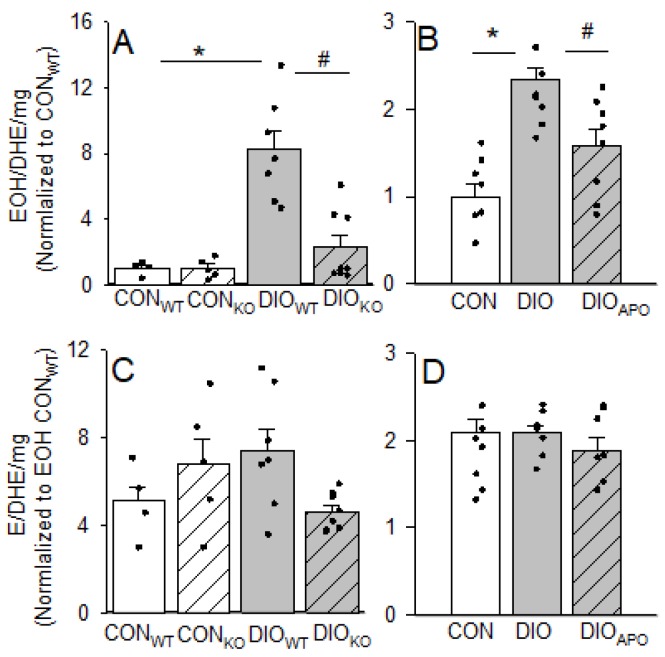
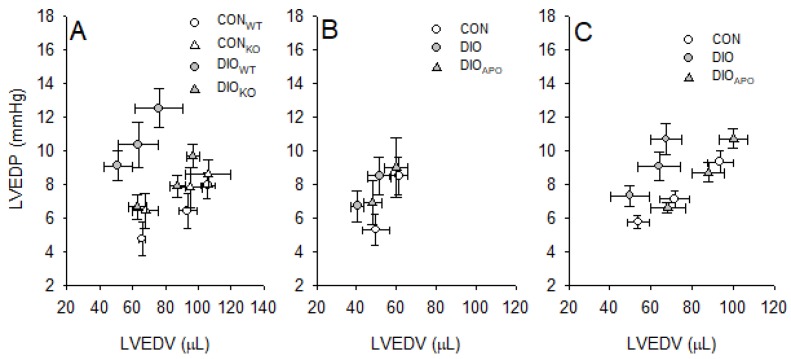
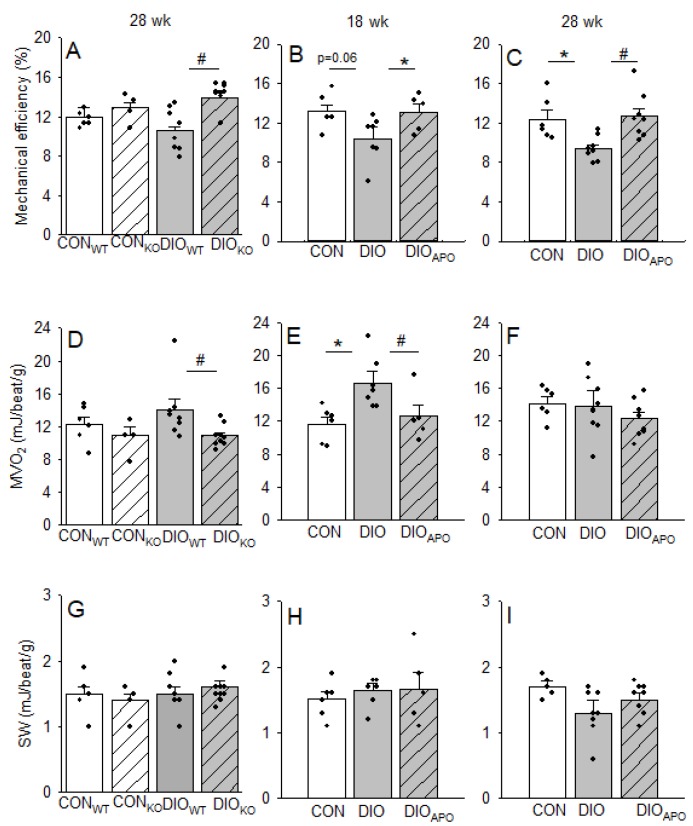
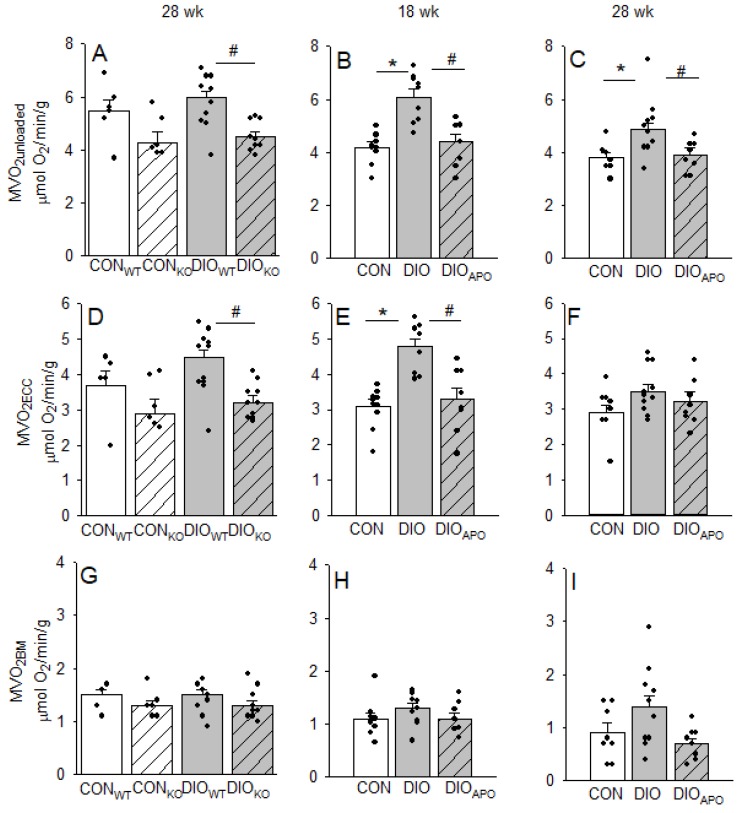
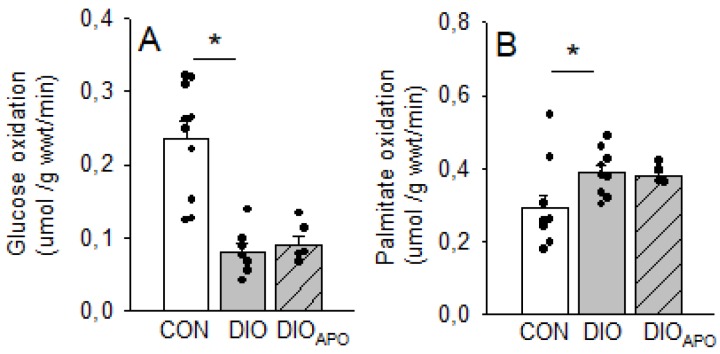
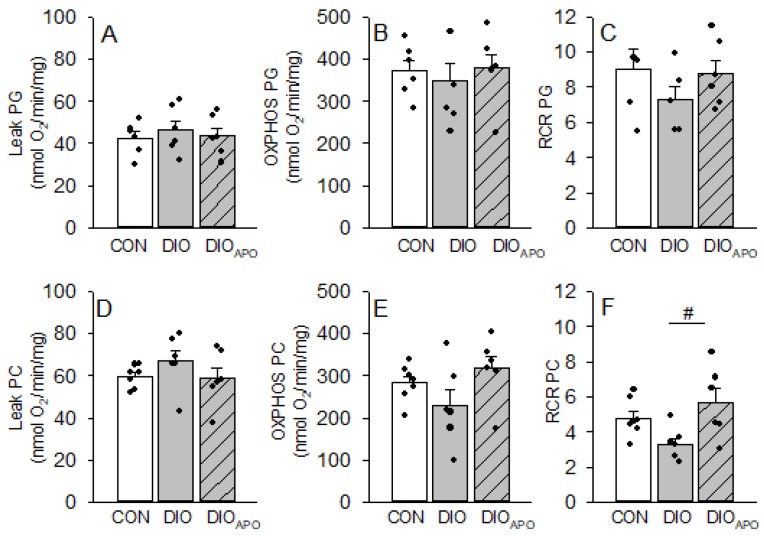
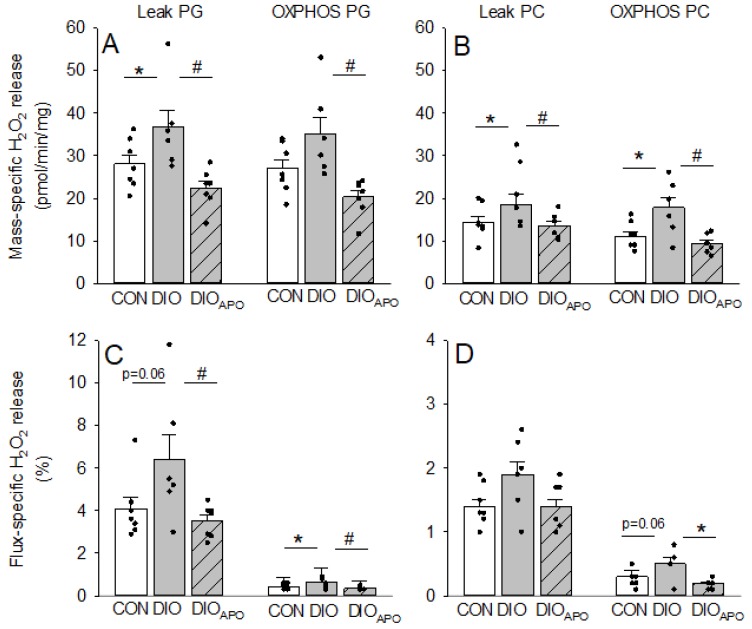
Obesity and diabetes are independent risk factors for cardiovascular diseases, and they are associated with the development of a specific cardiomyopathy with elevated myocardial oxygen consumption (MVO2) and impaired cardiac efficiency. Although the pathophysiology of this cardiomyopathy is multifactorial and complex, reactive oxygen species (ROS) may play an important role. One of the major ROS-generating enzymes in the cardiomyocytes is nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 2 (NOX2), and many potential systemic activators of NOX2 are elevated in obesity and diabetes. We hypothesized that NOX2 activity would influence cardiac energetics and/or the progression of ventricular dysfunction following obesity. Myocardial ROS content and mechanoenergetics were measured in the hearts from diet-induced-obese wild type (DIOWT) and global NOK2 knock-out mice (DIOKO) and in diet-induced obese C57BL/6J mice given normal water (DIO) or water supplemented with the NOX2-inhibitor apocynin (DIOAPO). Mitochondrial function and ROS production were also assessed in DIO and DIOAPO mice. This study demonstrated that ablation and pharmacological inhibition of NOX2 both improved mechanical efficiency and reduced MVO2 for non-mechanical cardiac work. Mitochondrial ROS production was also reduced following NOX2 inhibition, while cardiac mitochondrial function was not markedly altered by apocynin-treatment. Therefore, these results indicate a link between obesity-induced myocardial oxygen wasting, NOX2 activation, and mitochondrial ROS.
Keywords: NADPH-oxidase; ROS; cardiac efficiency; metabolism; myocardial oxygen consumption; obesity.
Conflict of interest statement
“The authors declare no conflict of interest.”
Figures
References
-
- Anderson E.J., Kypson A.P., Rodriguez E., Anderson C.A., Lehr E.J., Neufer P.D. Substrate-specific derangements in mitochondrial metabolism and redox balance in the atrium of the type 2 diabetic human heart. J. Am. Coll. Cardiol. 2009;54:1891–1898. doi: 10.1016/j.jacc.2009.07.031. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
