

Mammalian lectin arrays for screening host-microbe interactions
- PMID: 32094229
- PMCID: PMC7135977
- DOI: 10.1074/jbc.RA120.012783
Mammalian lectin arrays for screening host-microbe interactions
Abstract
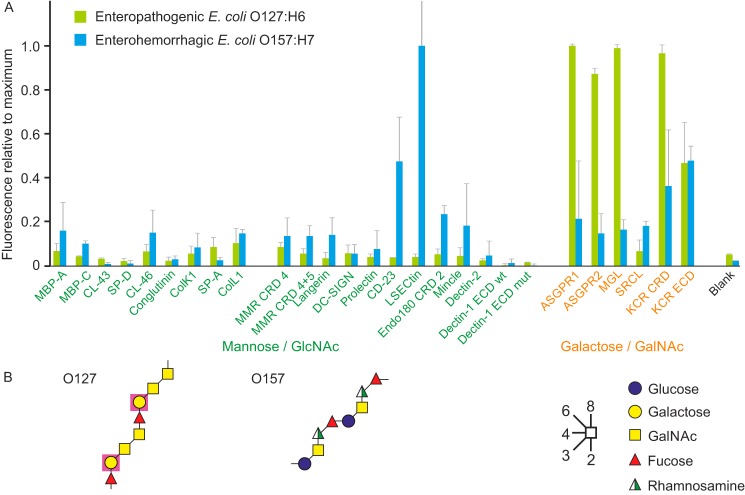
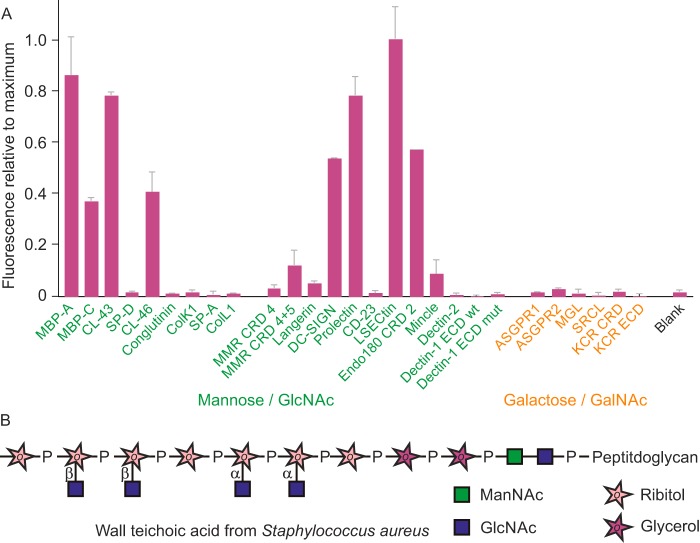
Many members of the C-type lectin family of glycan-binding receptors have been ascribed roles in the recognition of microorganisms and serve as key receptors in the innate immune response to pathogens. Other mammalian receptors have become targets through which pathogens enter target cells. These receptor roles have often been documented with binding studies involving individual pairs of receptors and microorganisms. To provide a systematic overview of interactions between microbes and the large complement of C-type lectins, here we developed a lectin array and suitable protocols for labeling of microbes that could be used to probe this array. The array contains C-type lectins from cow, chosen as a model organism of agricultural interest for which the relevant pathogen-receptor interactions have not been previously investigated in detail. Screening with yeast cells and various strains of both Gram-positive and -negative bacteria revealed distinct binding patterns, which in some cases could be explained by binding to lipopolysaccharides or capsular polysaccharides, but in other cases they suggested the presence of novel glycan targets on many of the microorganisms. These results are consistent with interactions previously ascribed to the receptors, but they also highlight binding to additional sugar targets that have not previously been recognized. Our findings indicate that mammalian lectin arrays represent unique discovery tools for identifying both novel ligands and new receptor functions.
Keywords: array screening; carbohydrate function; carbohydrate-binding protein; glycan-binding receptors; glycobiology; innate immune system; lectin; ligand binding; lipopolysaccharide (LPS); pathogen.
© 2020 Jégouzo et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures








References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
