

Oligomers of the lipodystrophy protein seipin may co-ordinate GPAT3 and AGPAT2 enzymes to facilitate adipocyte differentiation
- PMID: 32094408
- PMCID: PMC7039881
- DOI: 10.1038/s41598-020-59982-5
Oligomers of the lipodystrophy protein seipin may co-ordinate GPAT3 and AGPAT2 enzymes to facilitate adipocyte differentiation
Abstract
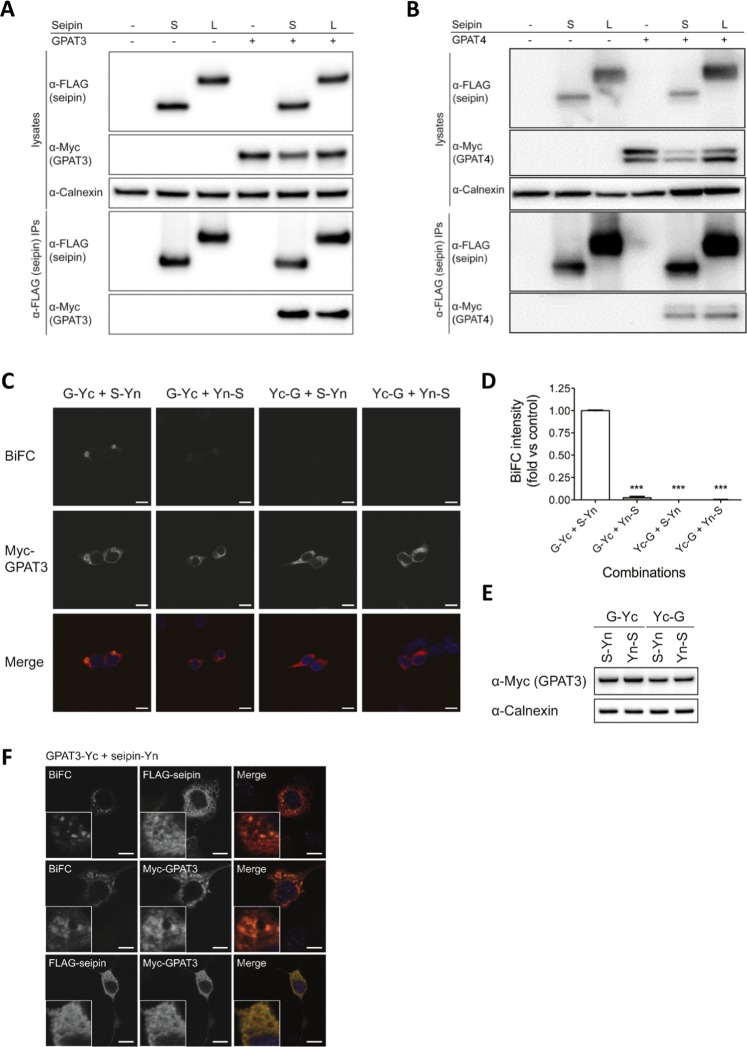
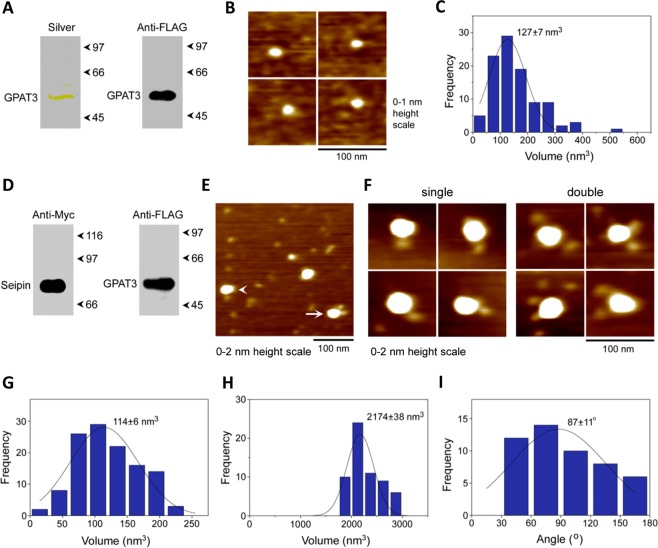
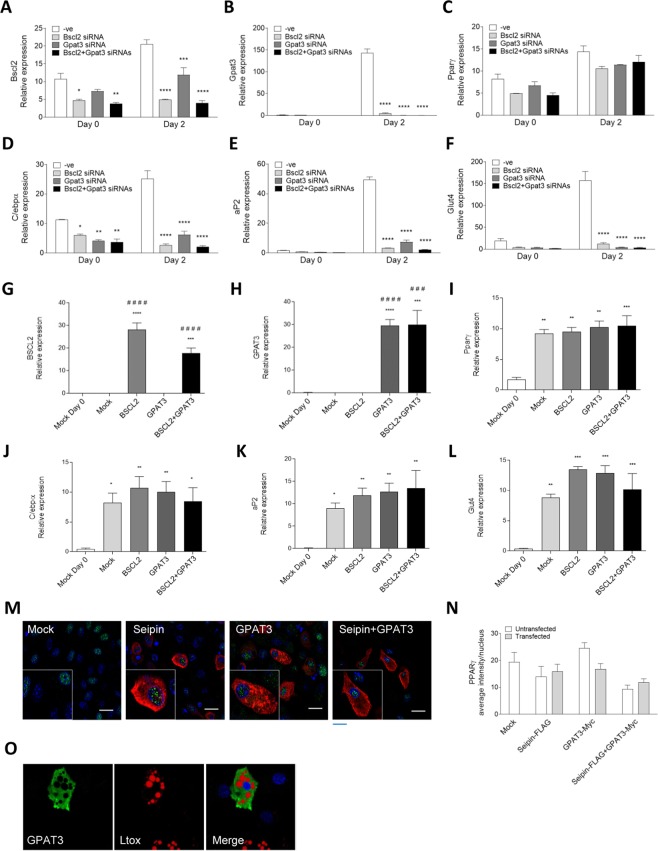
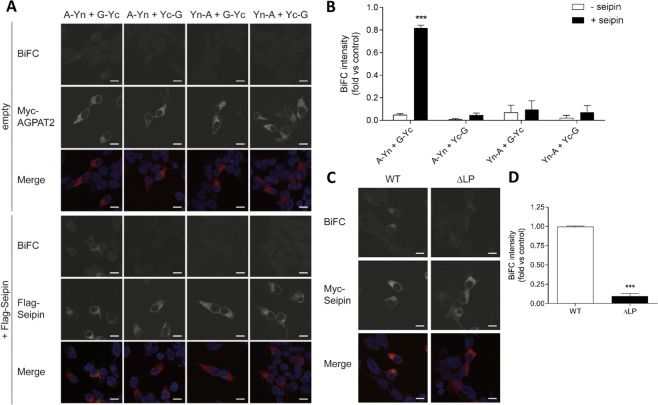
Seipin deficiency causes severe congenital generalized lipodystrophy (CGL) and metabolic disease. However, how seipin regulates adipocyte development and function remains incompletely understood. We previously showed that seipin acts as a scaffold protein for AGPAT2, whose disruption also causes CGL. More recently, seipin has been reported to promote adipogenesis by directly inhibiting GPAT3, leading to the suggestion that GPAT inhibitors could offer novel treatments for CGL. Here we investigated the interactions between seipin, GPAT3 and AGPAT2. We reveal that seipin and GPAT3 associate via direct interaction and that seipin can simultaneously bind GPAT3 and AGPAT2. Inhibiting the expression of seipin, AGPAT2 or GPAT3 led to impaired induction of early markers of adipocyte differentiation in cultured cells. However, consistent with normal adipose mass in GPAT3-null mice, GPAT3 inhibition did not prevent the formation of mature adipocytes. Nonetheless, loss of GPAT3 in seipin-deficient preadipocytes exacerbated the failure of adipogenesis in these cells. Thus, our data indicate that GPAT3 plays a modest positive role in adipogenesis and argue against the potential of GPAT inhibitors to rescue white adipose tissue mass in CGL2. Overall, our study reveals novel mechanistic insights regarding the molecular pathogenesis of severe lipodystrophy caused by mutations in either seipin or AGPAT2.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
