

Components of the endocytic and recycling trafficking pathways interfere with the integrity of the Legionella-containing vacuole
- PMID: 32096265
- PMCID: PMC7154685
- DOI: 10.1111/cmi.13151
Components of the endocytic and recycling trafficking pathways interfere with the integrity of the Legionella-containing vacuole
Abstract
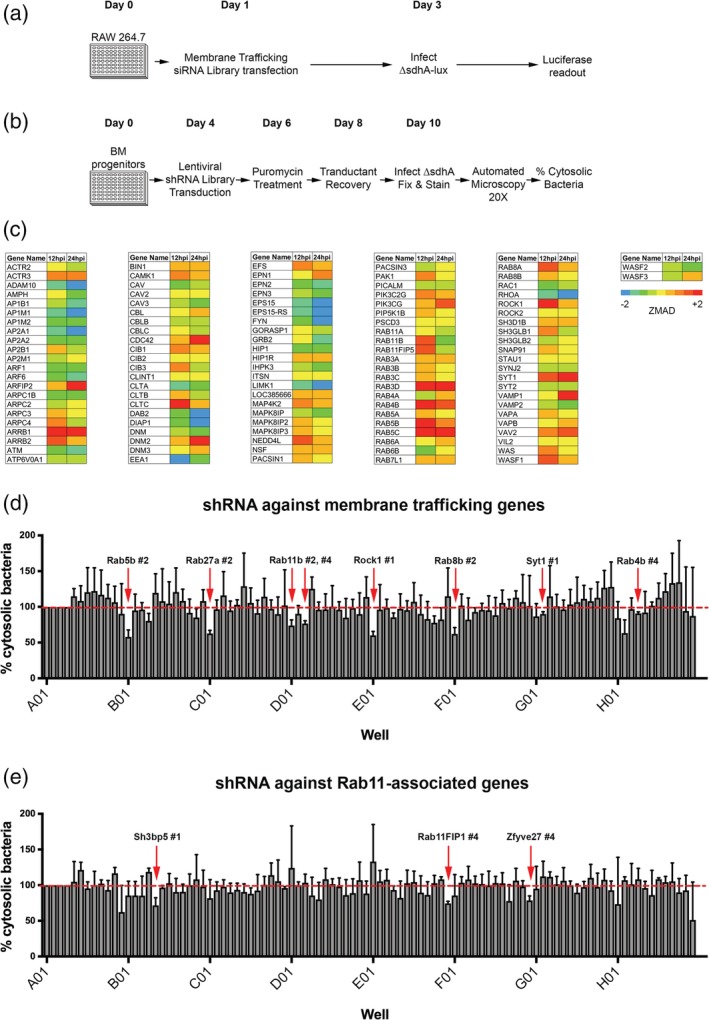
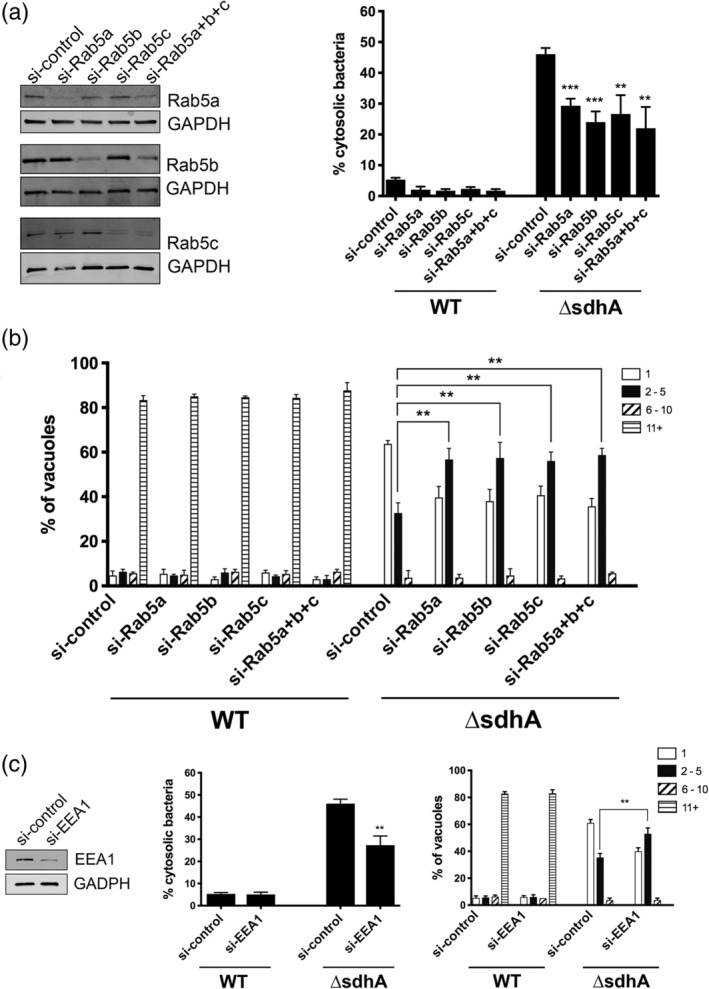
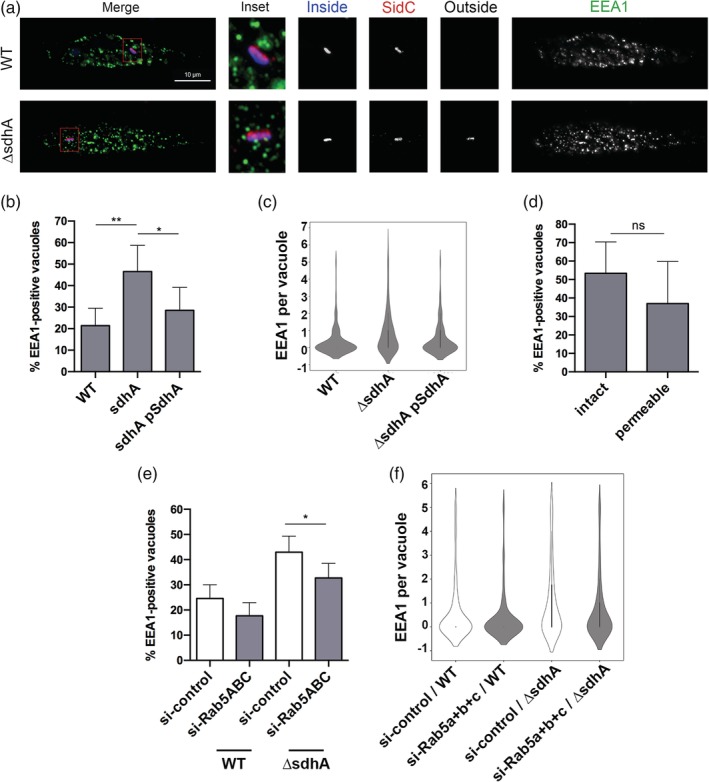
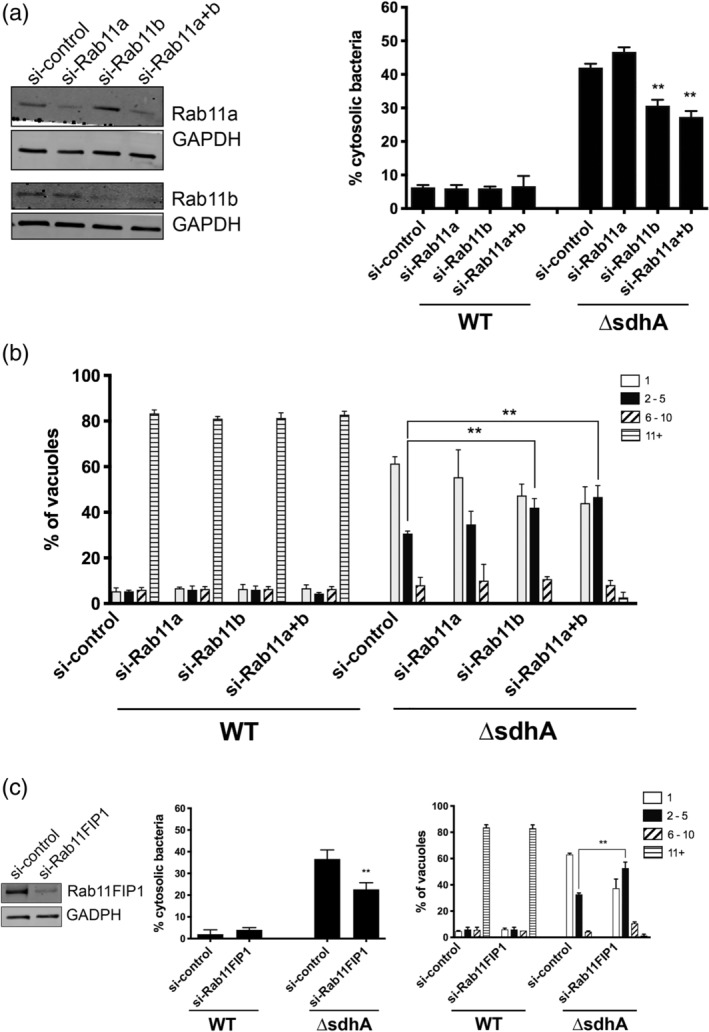
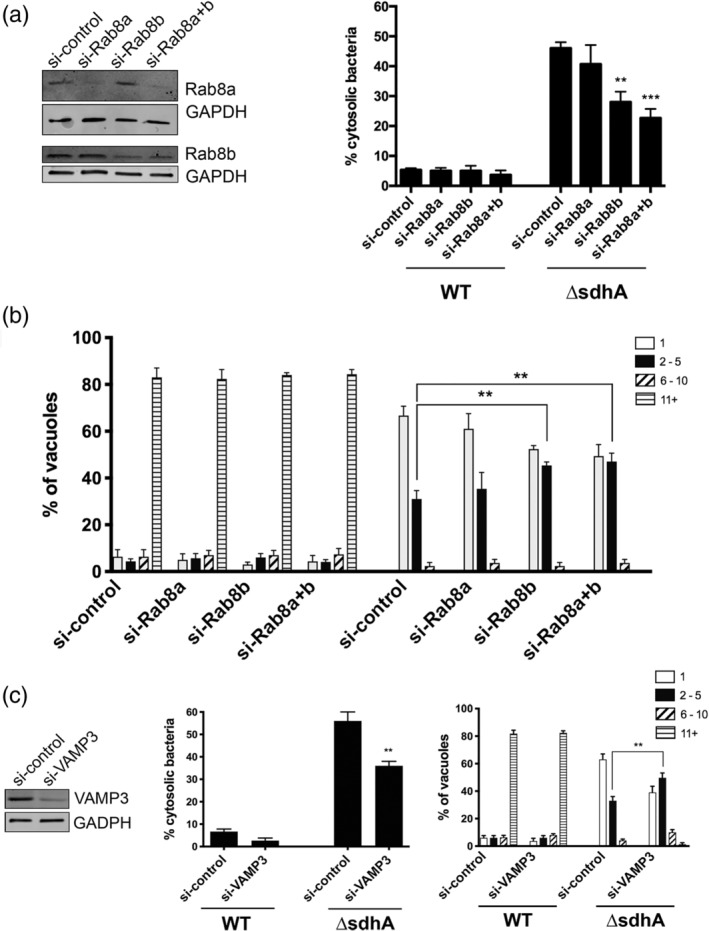
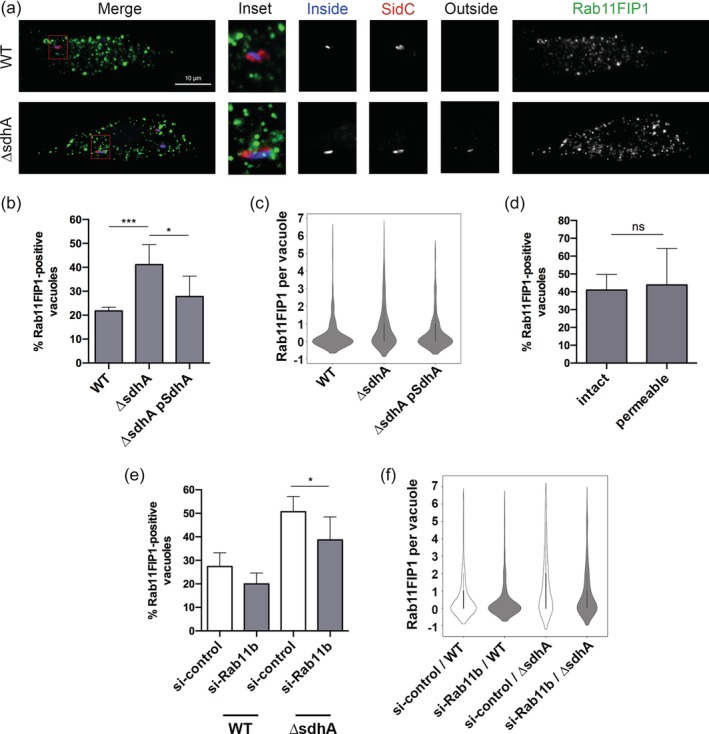
Legionella pneumophila requires the Dot/Icm translocation system to replicate in a vacuolar compartment within host cells. Strains lacking the translocated substrate SdhA form a permeable vacuole during residence in the host cell, exposing bacteria to the host cytoplasm. In primary macrophages, mutants are defective for intracellular growth, with a pyroptotic cell death response mounted due to bacterial exposure to the cytosol. To understand how SdhA maintains vacuole integrity during intracellular growth, we performed high-throughput RNAi screens against host membrane trafficking genes to identify factors that antagonise vacuole integrity in the absence of SdhA. Depletion of host proteins involved in endocytic uptake and recycling resulted in enhanced intracellular growth and lower levels of permeable vacuoles surrounding the ΔsdhA mutant. Of interest were three different Rab GTPases involved in these processes: Rab11b, Rab8b and Rab5 isoforms, that when depleted resulted in enhanced vacuole integrity surrounding the sdhA mutant. Proteins regulated by these Rabs are responsible for interfering with proper vacuole membrane maintenance, as depletion of the downstream effectors EEA1, Rab11FIP1, or VAMP3 rescued vacuole integrity and intracellular growth of the sdhA mutant. To test the model that specific vesicular components associated with these effectors could act to destabilise the replication vacuole, EEA1 and Rab11FIP1 showed increased density about the sdhA mutant vacuole compared with the wild type (WT) vacuole. Depletion of Rab5 isoforms or Rab11b reduced this aberrant redistribution. These findings are consistent with SdhA interfering with both endocytic and recycling membrane trafficking events that act to destabilise vacuole integrity during infection.
Keywords: Legionella; RNAi; Rab proteins; intracellular growth; vesicle trafficking.
© 2020 The Authors. Cellular Microbiology published by John Wiley & Sons Ltd.
Figures
References
-
- Banerjee, M. , Joshi, S. , Zhang, J. , Moncman, C. L. , Yadav, S. , Bouchard, B. A. , … Whiteheart, S. W. (2017). Cellubrevin/vesicle‐associated membrane protein‐3‐mediated endocytosis and trafficking regulate platelet functions. Blood, 130(26), 2872–2883. 10.1182/blood-2017-02-768176 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
