

FLIP(L): the pseudo-caspase
- PMID: 32096279
- PMCID: PMC7586951
- DOI: 10.1111/febs.15260
FLIP(L): the pseudo-caspase
Abstract
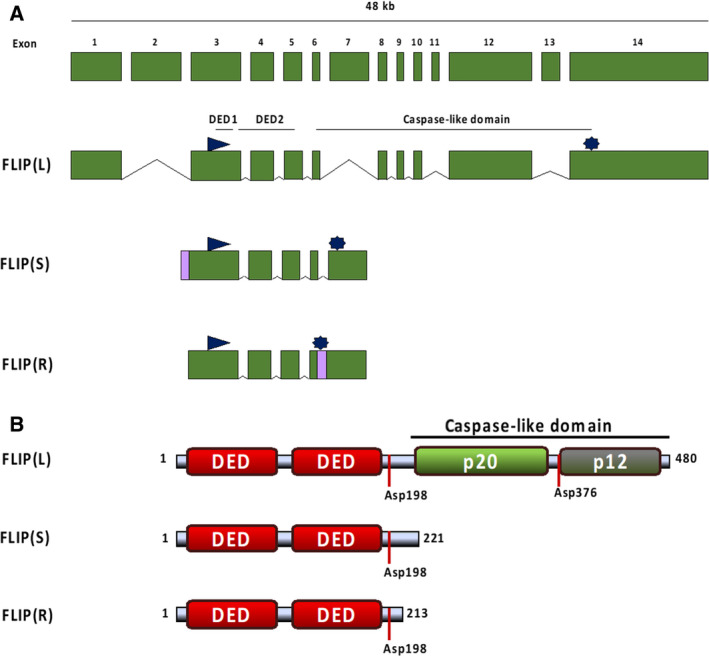
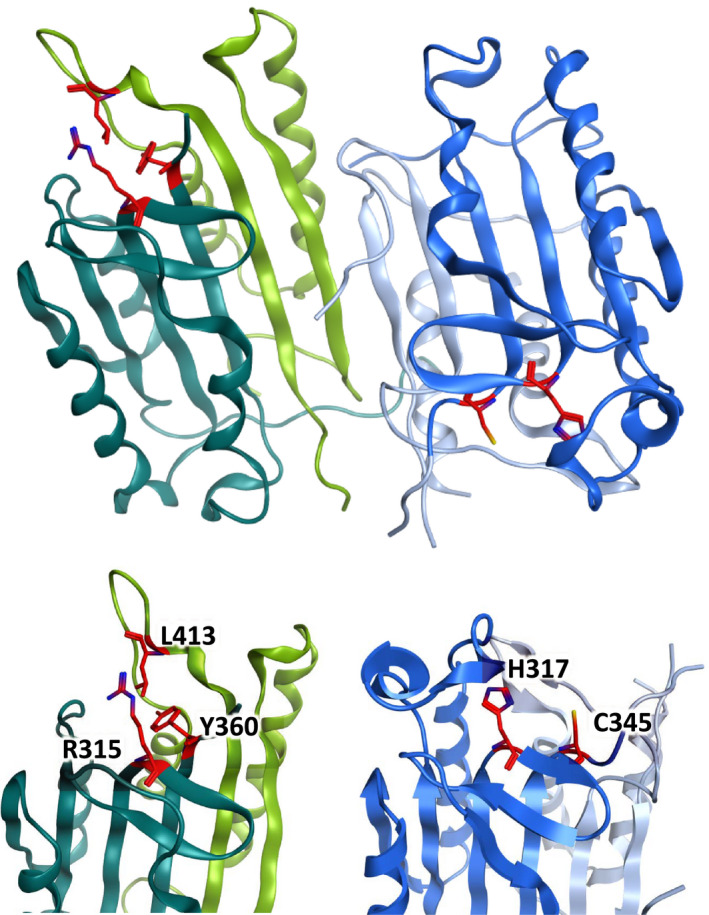
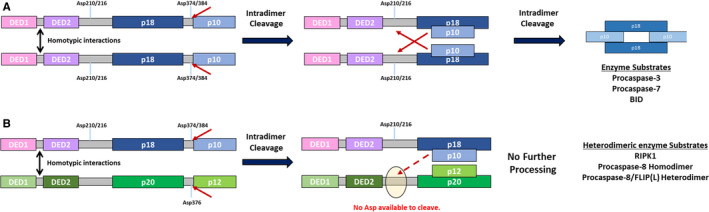
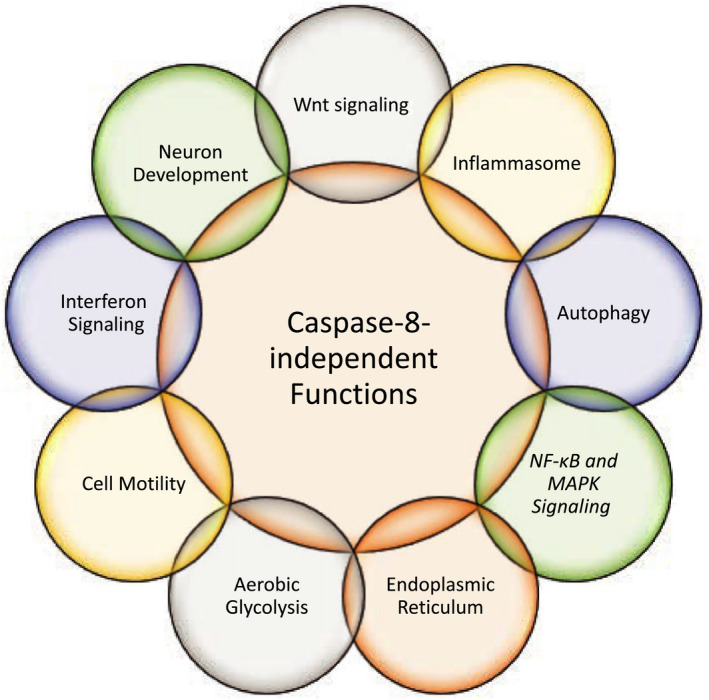
Possessing structural homology with their active enzyme counterparts but lacking catalytic activity, pseudoenzymes have been identified for all major enzyme groups. Caspases are a family of cysteine-dependent aspartate-directed proteases that play essential roles in regulating cell death and inflammation. Here, we discuss the only human pseudo-caspase, FLIP(L), a paralog of the apoptosis-initiating caspases, caspase-8 and caspase-10. FLIP(L) has been shown to play a key role in regulating the processing and activity of caspase-8, thereby modulating apoptotic signaling mediated by death receptors (such as TRAIL-R1/R2), TNF receptor-1 (TNFR1), and Toll-like receptors. In this review, these canonical roles of FLIP(L) are discussed. Additionally, a range of nonclassical pseudoenzyme roles are described, in which FLIP(L) functions independently of caspase-8. These nonclassical pseudoenzyme functions enable FLIP(L) to play key roles in the regulation of a wide range of biological processes beyond its canonical roles as a modulator of cell death.
Keywords: DISC; FLIP(L); apoptosis; autophagy; caspase; necroptosis; pseudo-caspase; pseudoenzymes.
© 2020 The Authors. The FEBS Journal published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Murphy JM, Mace PD & Eyers PA (2017) Live and let die: insights into pseudoenzyme mechanisms from structure. Curr Opin Struct Biol 47, 95–104. - PubMed
-
- Ribeiro AJM, Das S, Dawson N, Zaru R, Orchard S, Thornton JM, Orengo C, Zeqiraj E, Murphy JM & Eyers PA (2019) Emerging concepts in pseudoenzyme classification, evolution, and signaling. Sci Signal 12, eaat9797. - PubMed
-
- Jacobsen AV & Murphy JM (2017) The secret life of kinases: insights into non‐catalytic signalling functions from pseudokinases. Biochem Soc Trans 45, 665–681. - PubMed
-
- Jeffery CJ (2019) The demise of catalysis, but new functions arise: pseudoenzymes as the phoenixes of the protein world. Biochem Soc Trans 47, 371–379. - PubMed
-
- Adrain C & Freeman M (2012) New lives for old: evolution of pseudoenzyme function illustrated by iRhoms. Nat Rev Mol Cell Biol 13, 489–498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
