

The Plasmodium falciparum Artemisinin Susceptibility-Associated AP-2 Adaptin μ Subunit is Clathrin Independent and Essential for Schizont Maturation
- PMID: 32098816
- PMCID: PMC7042695
- DOI: 10.1128/mBio.02918-19
The Plasmodium falciparum Artemisinin Susceptibility-Associated AP-2 Adaptin μ Subunit is Clathrin Independent and Essential for Schizont Maturation
Abstract
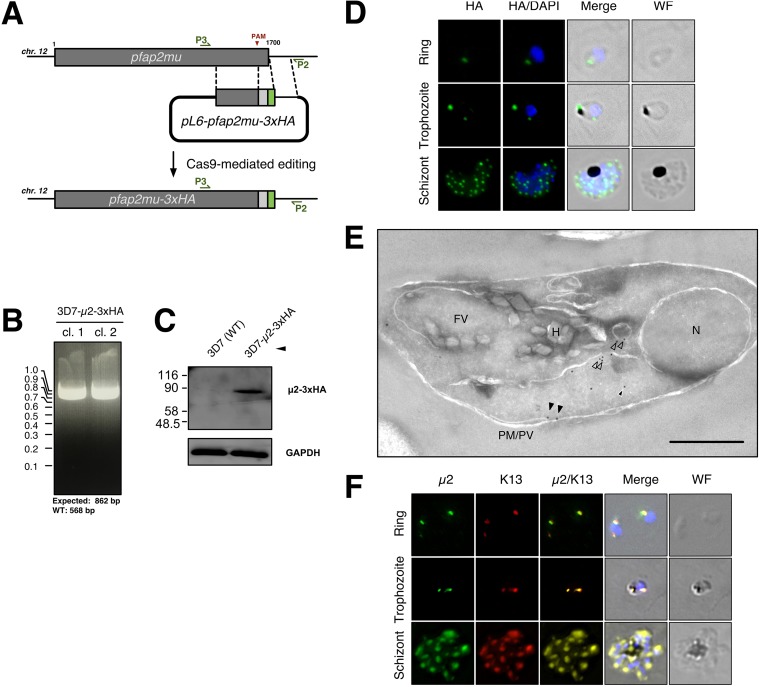
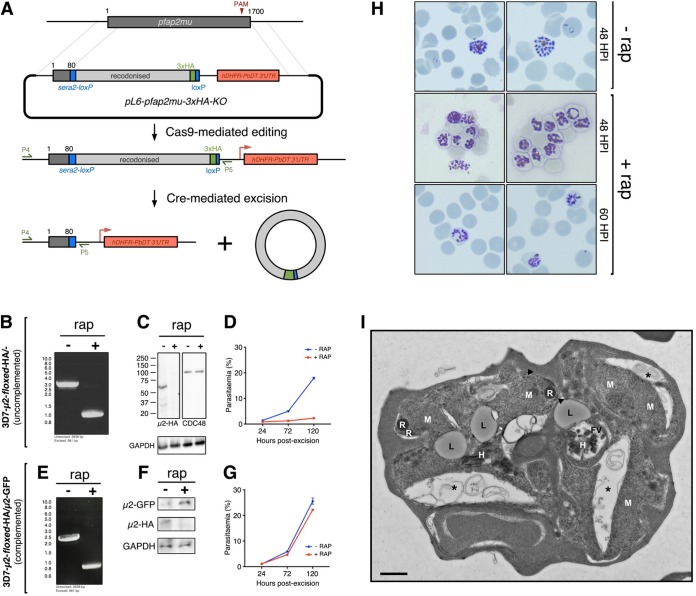
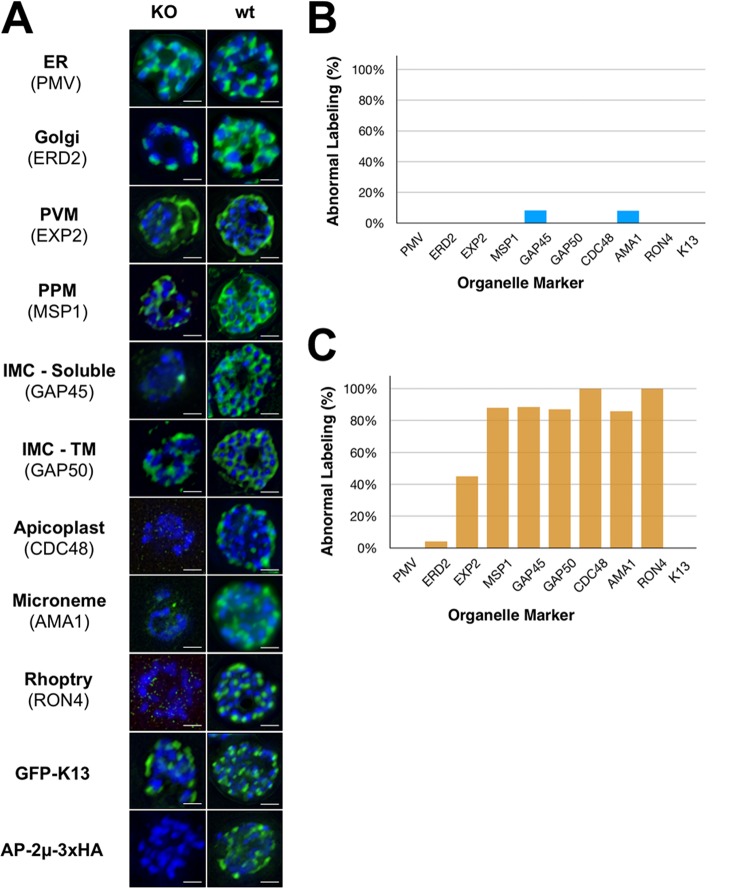
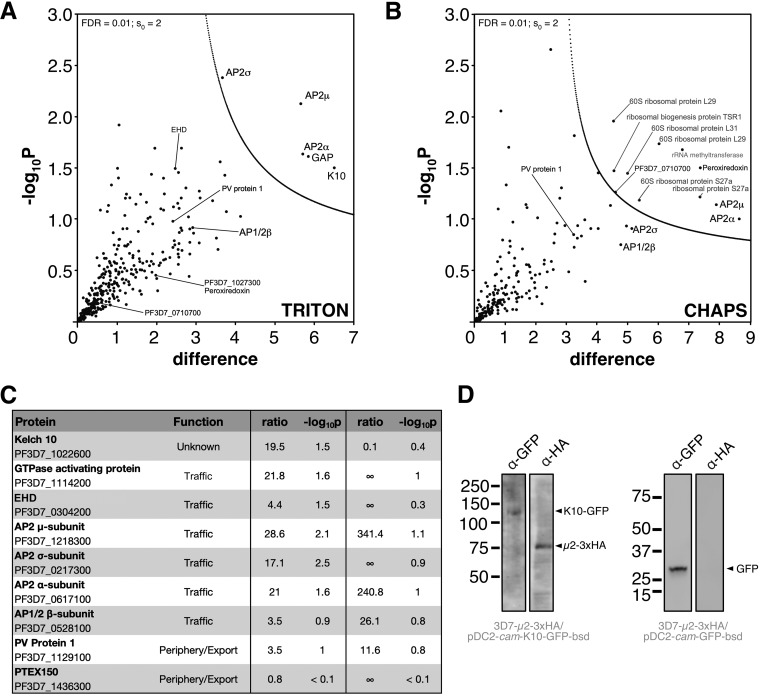
The efficacy of current antimalarial drugs is threatened by reduced susceptibility of Plasmodium falciparum to artemisinin, associated with mutations in pfkelch13 Another gene with variants known to modulate the response to artemisinin encodes the μ subunit of the AP-2 adaptin trafficking complex. To elucidate the cellular role of AP-2μ in P. falciparum, we performed a conditional gene knockout, which severely disrupted schizont organization and maturation, leading to mislocalization of key merozoite proteins. AP-2μ is thus essential for blood-stage replication. We generated transgenic P. falciparum parasites expressing hemagglutinin-tagged AP-2μ and examined cellular localization by fluorescence and electron microscopy. Together with mass spectrometry analysis of coimmunoprecipitating proteins, these studies identified AP-2μ-interacting partners, including other AP-2 subunits, the K10 kelch-domain protein, and PfEHD, an effector of endocytosis and lipid mobilization, but no evidence was found of interaction with clathrin, the expected coat protein for AP-2 vesicles. In reverse immunoprecipitation experiments with a clathrin nanobody, other heterotetrameric AP-complexes were shown to interact with clathrin, but AP-2 complex subunits were absent.IMPORTANCE We examine in detail the AP-2 adaptin complex from the malaria parasite Plasmodium falciparum In most studied organisms, AP-2 is involved in bringing material into the cell from outside, a process called endocytosis. Previous work shows that changes to the μ subunit of AP-2 can contribute to drug resistance. Our experiments show that AP-2 is essential for parasite development in blood but does not have any role in clathrin-mediated endocytosis. This suggests that a specialized function for AP-2 has developed in malaria parasites, and this may be important for understanding its impact on drug resistance.
Keywords: Plasmodium falciparum; adaptin trafficking complex; adaptor proteins; artemisinin susceptibility; endocytosis; malaria.
Copyright © 2020 Henrici et al.
Figures
References
-
- World Health Organization. 2017. World malaria report. World Health Organization, Geneva, Switzerland.
-
- Thanh NV, Thuy-Nhien N, Tuyen NTK, Tong NT, Nha-Ca NT, Dong LT, Quang HH, Farrar J, Thwaites G, White NJ, Wolbers M, Hien TT. 2017. Rapid decline in the susceptibility of Plasmodium falciparum to dihydroartemisinin-piperaquine in the south of Vietnam. Malar J 16:27. doi: 10.1186/s12936-017-1680-8. - DOI - PMC - PubMed
-
- Ariey F, Witkowski B, Amaratunga C, Beghain J, Langlois A-C, Khim N, Kim S, Duru V, Bouchier C, Ma L, Lim P, Leang R, Duong S, Sreng S, Suon S, Chuor CM, Bout DM, Ménard S, Rogers WO, Genton B, Fandeur T, Miotto O, Ringwald P, Le Bras J, Berry A, Barale J-C, Fairhurst RM, Benoit-Vical F, Mercereau-Puijalon O, Ménard D. 2014. A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 505:50–55. doi: 10.1038/nature12876. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
