

Photoperiod and temperature separately regulate nymphal development through JH and insulin/TOR signaling pathways in an insect
- PMID: 32098850
- PMCID: PMC7071924
- DOI: 10.1073/pnas.1922747117
Photoperiod and temperature separately regulate nymphal development through JH and insulin/TOR signaling pathways in an insect
Abstract
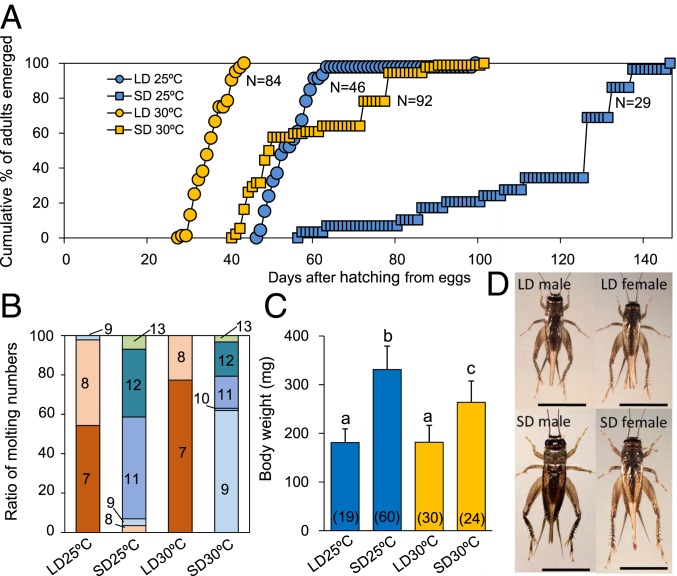
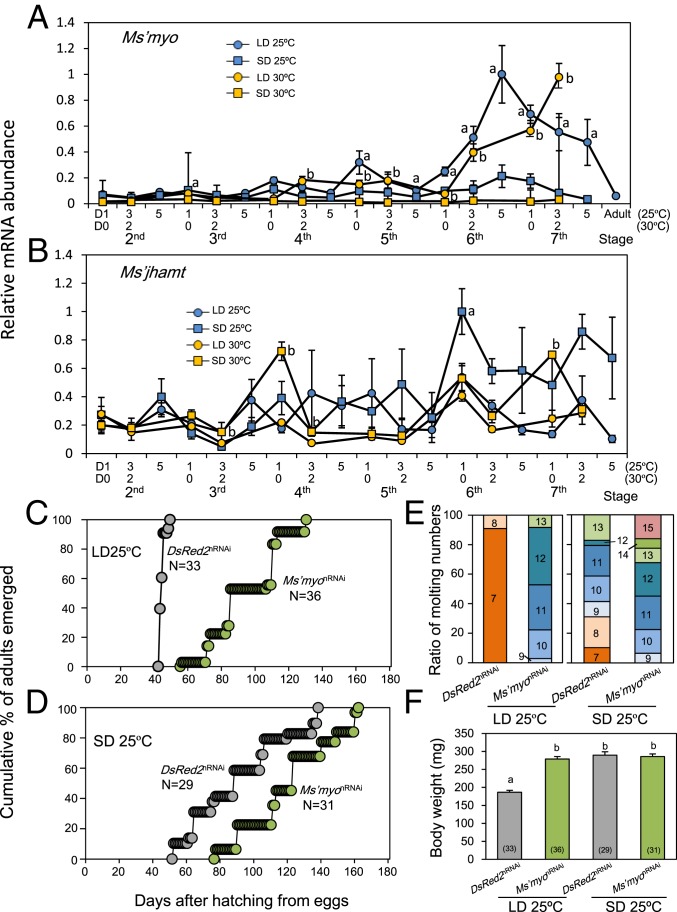
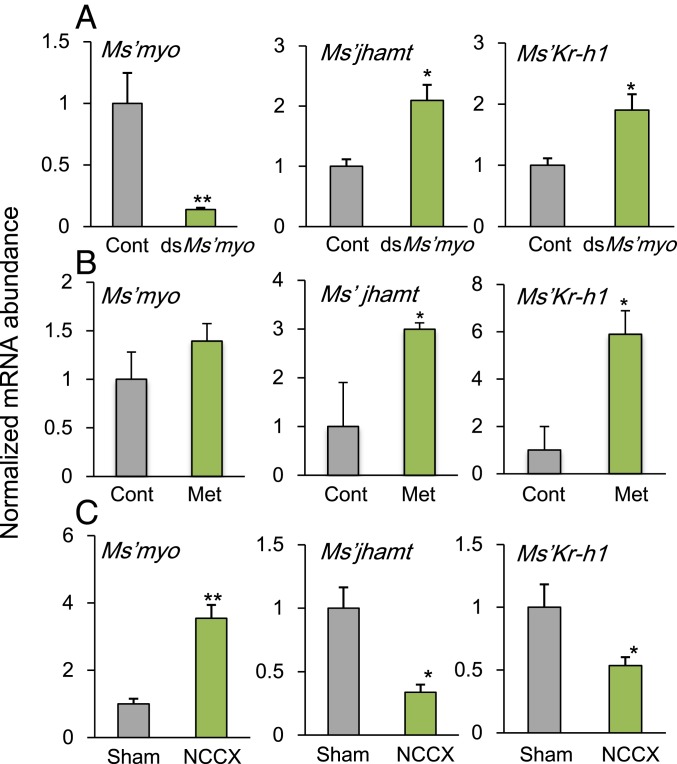
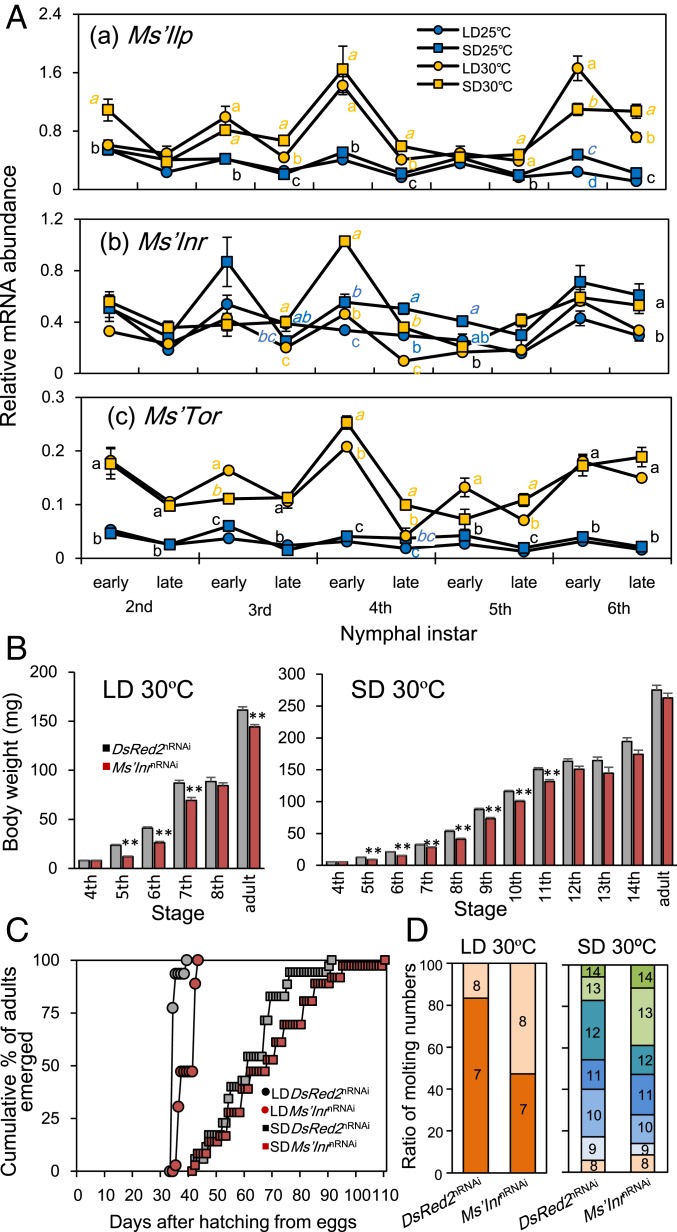
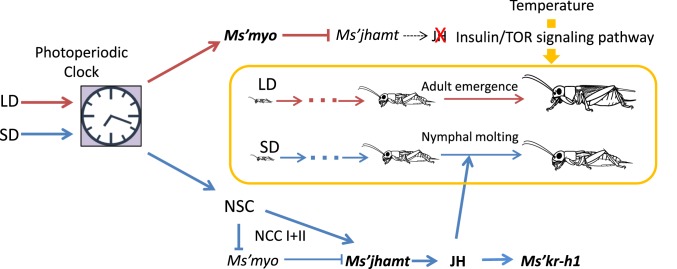
Insects living in the temperate zone enter a physiological state of arrested or slowed development to overcome an adverse season, such as winter. Developmental arrest, called diapause, occurs at a species-specific developmental stage, and embryonic and pupal diapauses have been extensively studied in mostly holometabolous insects. Some other insects overwinter in the nymphal stage with slow growth for which the mechanism is poorly understood. Here, we show that this nymphal period of slow growth is regulated by temperature and photoperiod through separate pathways in the cricket Modicogryllus siamensis The former regulates the growth rate, at least in part, through the insulin / target of rapamycin (TOR) signaling pathway. Lower temperature down-regulates the expression of insulin-like peptide (Ms'Ilp) and Target of rapamycin (Ms'Tor) genes to slow down the growth rate without affecting the number of molts. The latter regulates the number of molts independent of temperature. Short days increase the number of molts through activation of the juvenile hormone (JH) pathway and down-regulation of myoglianin (Ms'myo), a member of the TGFβ family, which induces adult metamorphosis. In contrast, long days regulate Ms'myo expression to increase during the fifth to sixth instar to initiate adult metamorphosis. When Ms'myo expression is suppressed, juvenile hormone O-methyl transferase (Ms'jhamt) was up-regulated and increased molts to prolong the nymphal period even under long-day conditions. The present findings suggested that the photoperiod regulated Ms'myo, and the JH signaling pathway and the temperature-controlled insulin/TOR pathway cooperated to regulate nymphal development for overwintering to achieve seasonal adaptation of the life cycle in M. siamensis.
Keywords: insulin/TOR signaling pathway; myo; nymphal development; photoperiod; temperature.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Saunders D. S., Steel C. G. H., Vafopoulou X., Lewis R. D., Insect Clocks (Elsevier, Amsterdam, ed. 3, 2002), p. 560.
-
- Beck S. D., Insect Photoperiodism (Academic Press, New York, ed. 2, 1980), p. 387.
-
- Saunders D. S., Insect photoperiodism: Seeing the light. Physiol. Entomol. 37, 207–218 (2012).
-
- Stehlík J., Závodská R., Shimada K., Sauman I., Kostál V., Photoperiodic induction of diapause requires regulated transcription of timeless in the larval brain of Chymomyza costata. J. Biol. Rhythms 23, 129–139 (2008). - PubMed
-
- Sakamoto T., Uryu O., Tomioka K., The clock gene period plays an essential role in photoperiodic control of nymphal development in the cricket Modicogryllus siamensis. J. Biol. Rhythms 24, 379–390 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
