

New opportunities and insights into Papaver self-incompatibility by imaging engineered Arabidopsis pollen
- PMID: 32100005
- PMCID: PMC7178406
- DOI: 10.1093/jxb/eraa092
New opportunities and insights into Papaver self-incompatibility by imaging engineered Arabidopsis pollen
Abstract
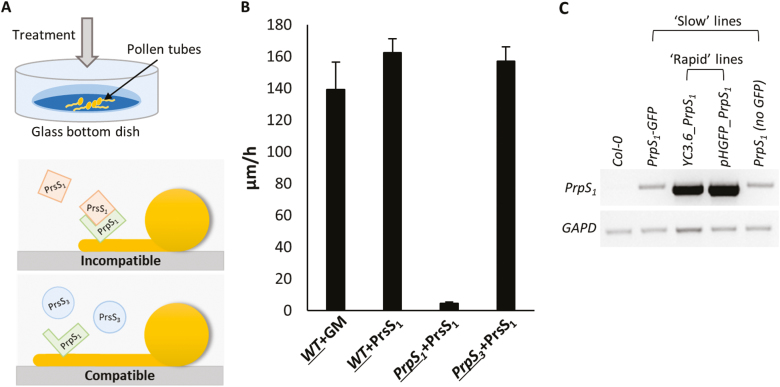
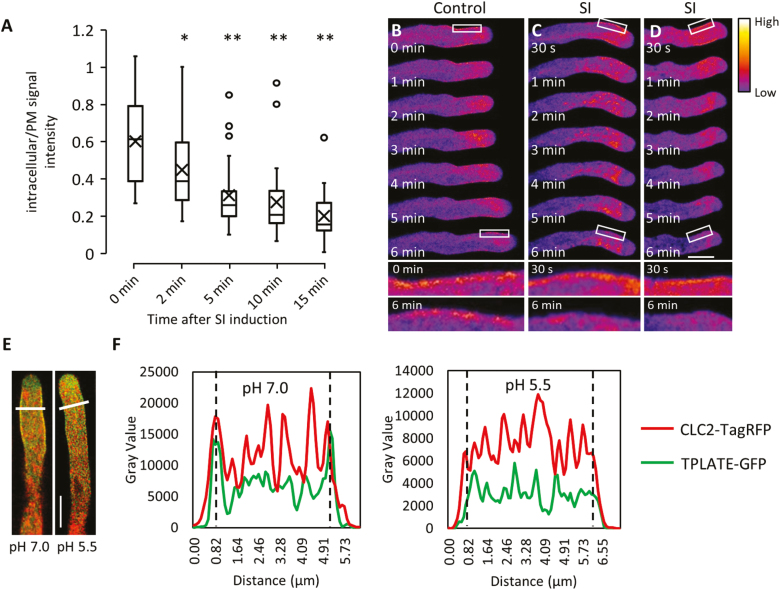
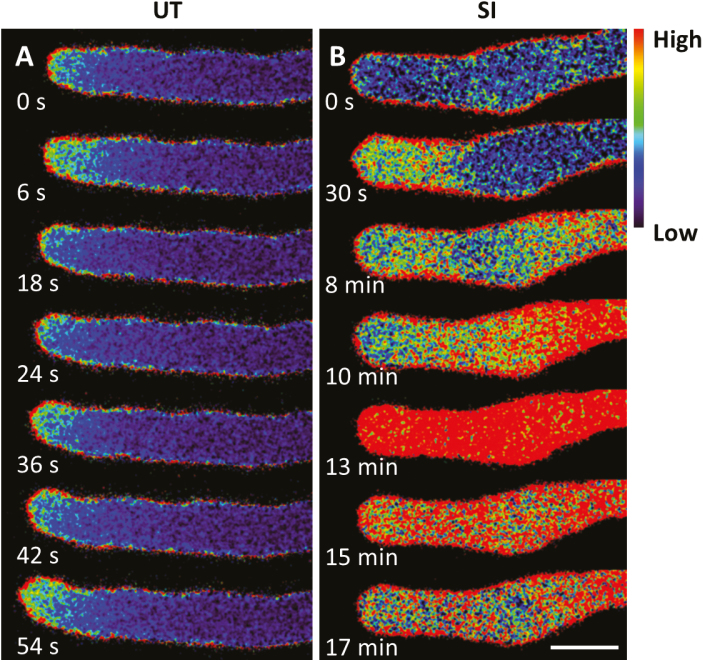
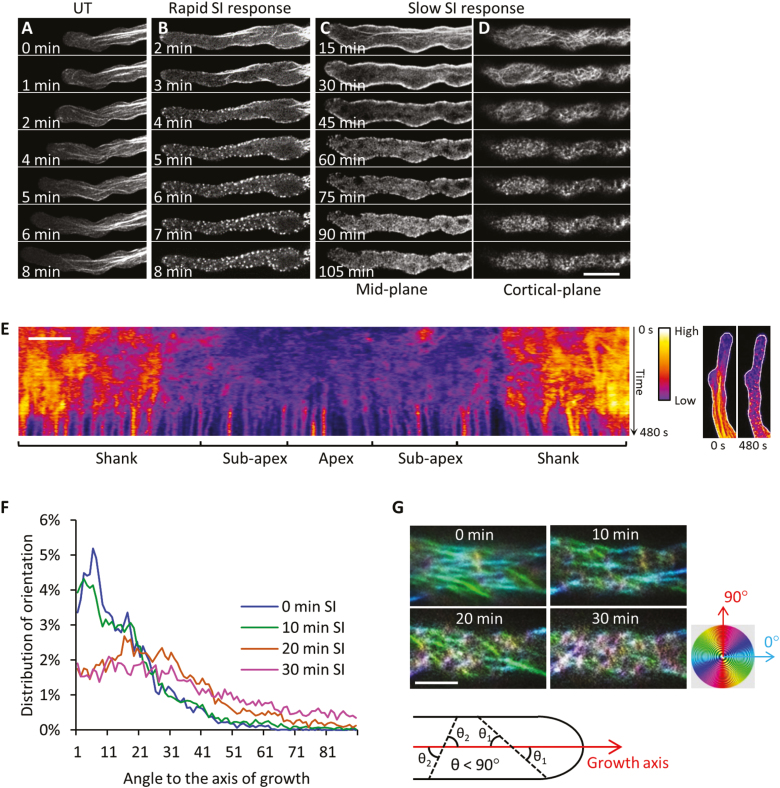
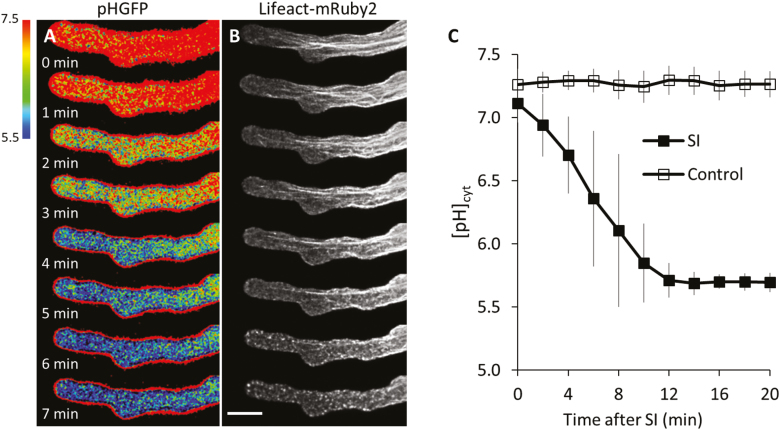
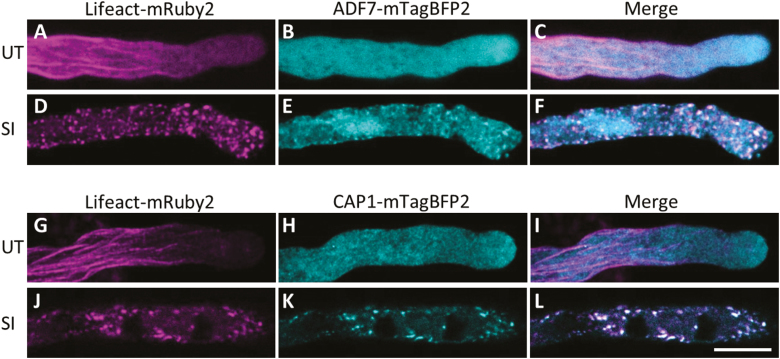
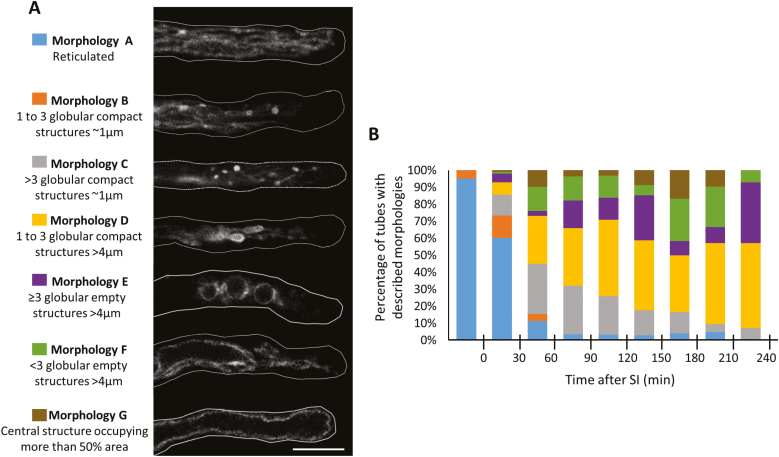
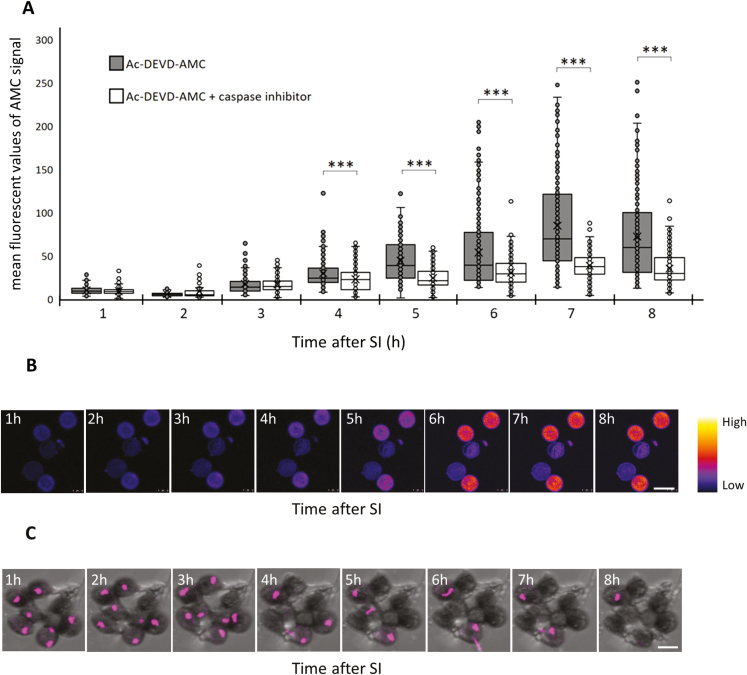
Pollen tube growth is essential for plant reproduction. Their rapid extension using polarized tip growth provides an exciting system for studying this specialized type of growth. Self-incompatibility (SI) is a genetically controlled mechanism to prevent self-fertilization. Mechanistically, one of the best-studied SI systems is that of Papaver rhoeas (poppy). This utilizes two S-determinants: stigma-expressed PrsS and pollen-expressed PrpS. Interaction of cognate PrpS-PrsS triggers a signalling network, causing rapid growth arrest and programmed cell death (PCD) in incompatible pollen. We previously demonstrated that transgenic Arabidopsis thaliana pollen expressing PrpS-green fluorescent protein (GFP) can respond to Papaver PrsS with remarkably similar responses to those observed in incompatible Papaver pollen. Here we describe recent advances using these transgenic plants combined with genetically encoded fluorescent probes to monitor SI-induced cellular alterations, including cytosolic calcium, pH, the actin cytoskeleton, clathrin-mediated endocytosis (CME), and the vacuole. This approach has allowed us to study the SI response in depth, using multiparameter live-cell imaging approaches that were not possible in Papaver. This lays the foundations for new opportunities to elucidate key mechanisms involved in SI. Here we establish that CME is disrupted in self-incompatible pollen. Moreover, we reveal new detailed information about F-actin remodelling in pollen tubes after SI.
Keywords: Actin; actin-binding proteins (ABPs); calcium; endocytosis; fluorescent probes; live-cell imaging; pH; pollen tube growth; programmed cell death (PCD); self-incompatibility (SI).
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Barnanas B, Fridvalszky L. 1984. Adhesion and germination of differently treated maize pollen grains on the stigma. Acta Botanica Hungarica 30, 329–332.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
