

Ciliary Rab28 and the BBSome negatively regulate extracellular vesicle shedding
- PMID: 32101165
- PMCID: PMC7043889
- DOI: 10.7554/eLife.50580
Ciliary Rab28 and the BBSome negatively regulate extracellular vesicle shedding
Abstract
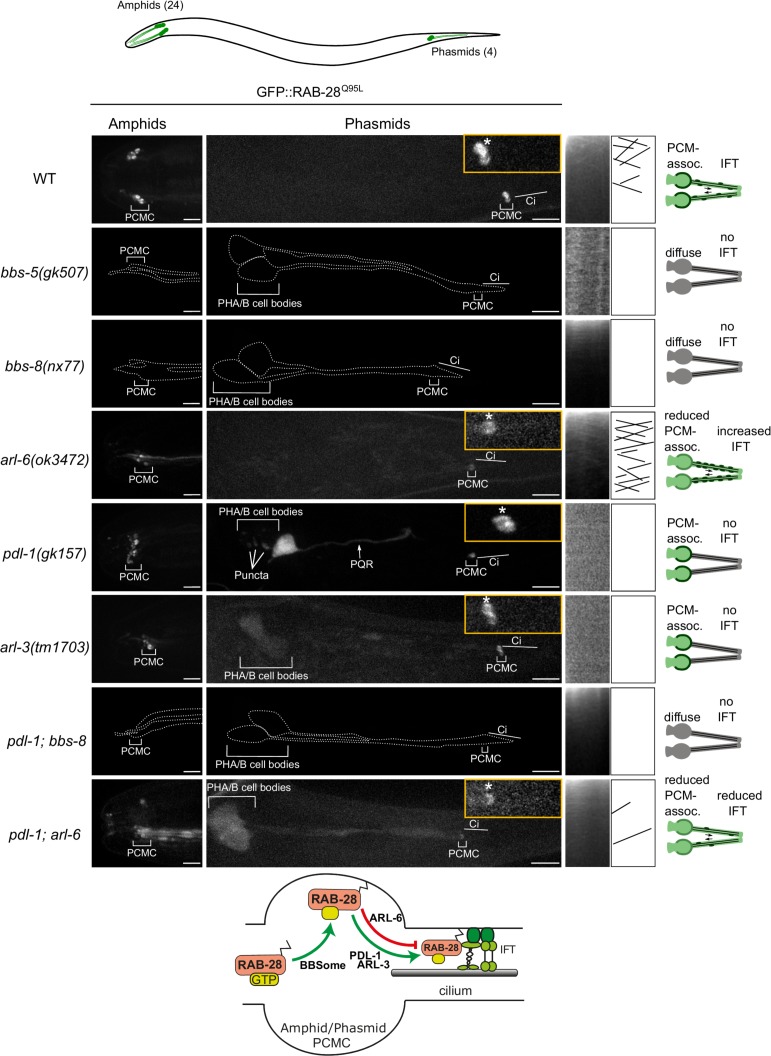
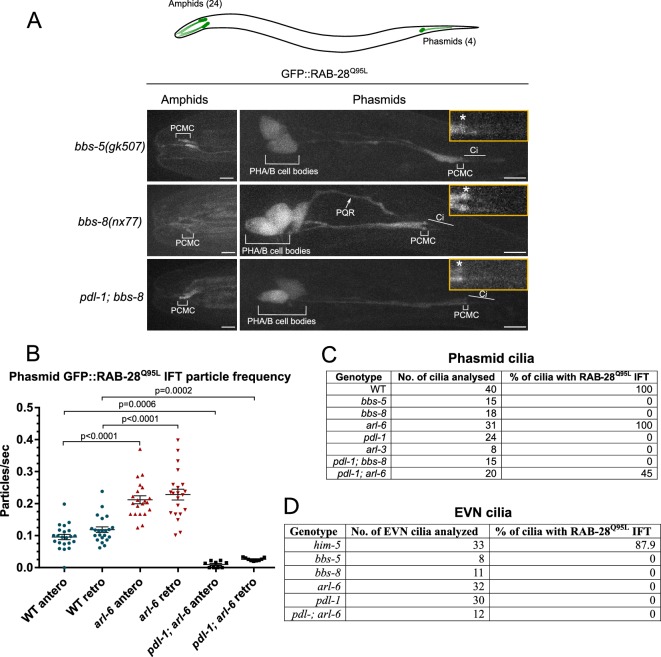
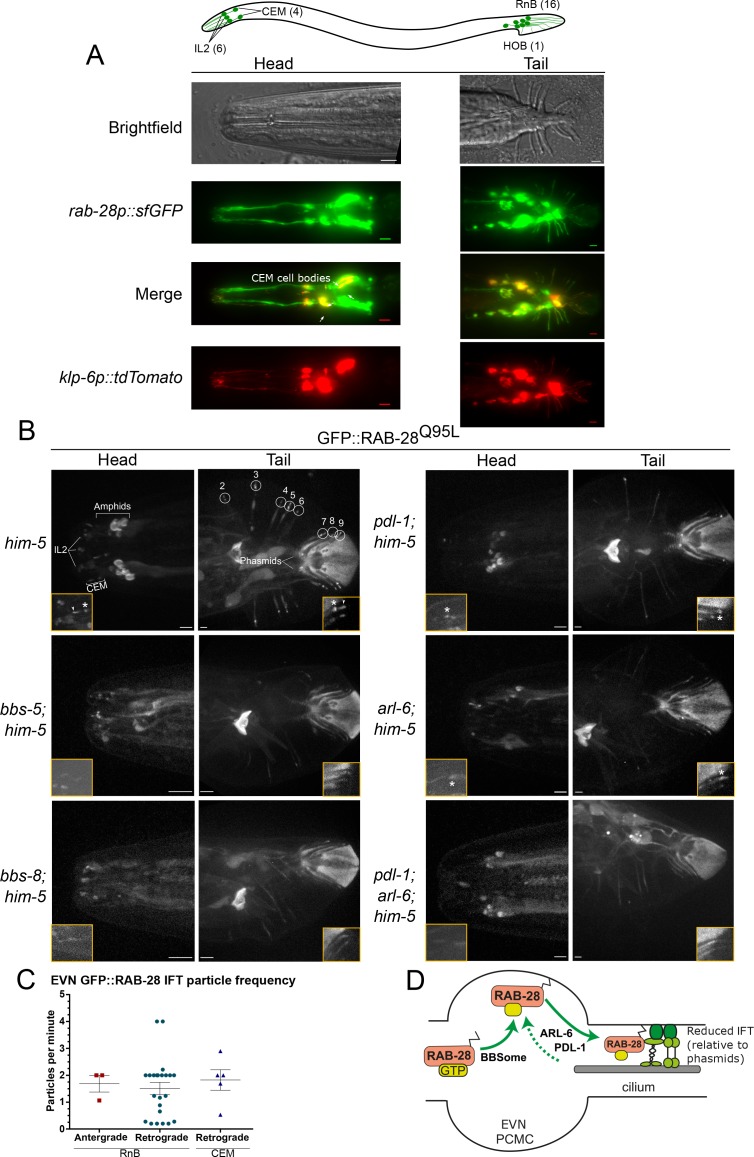
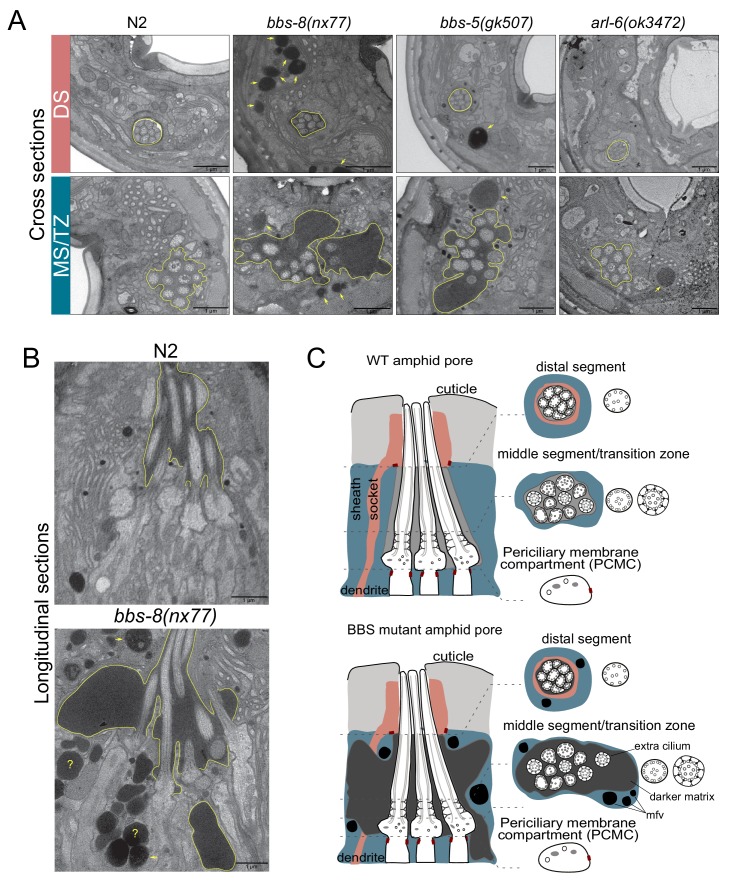
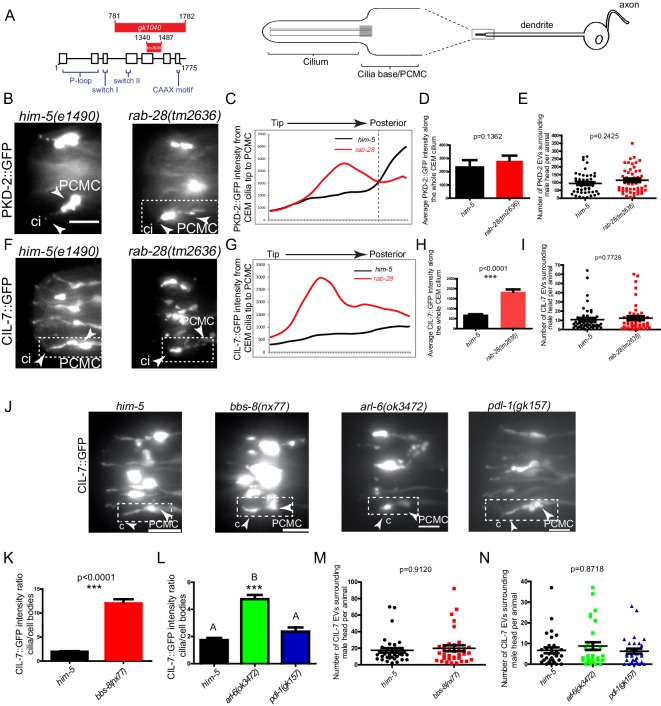
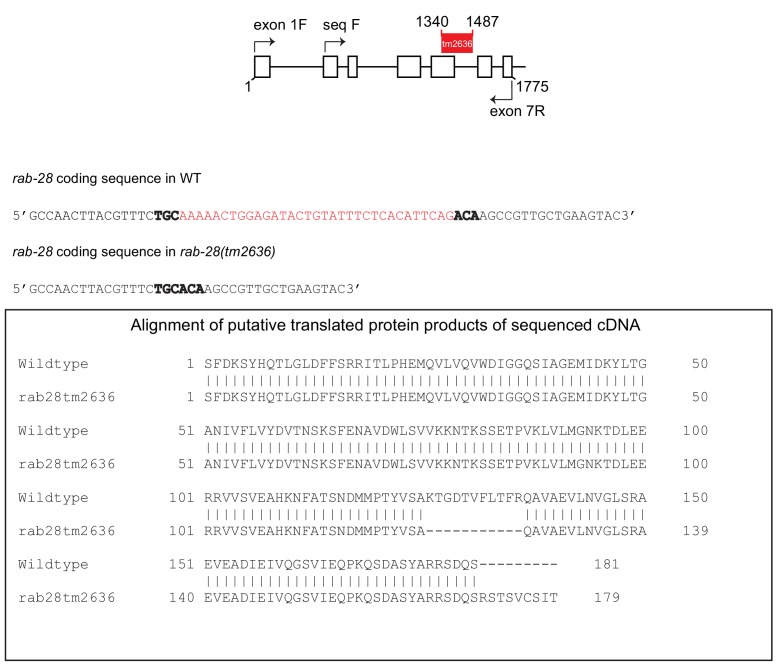
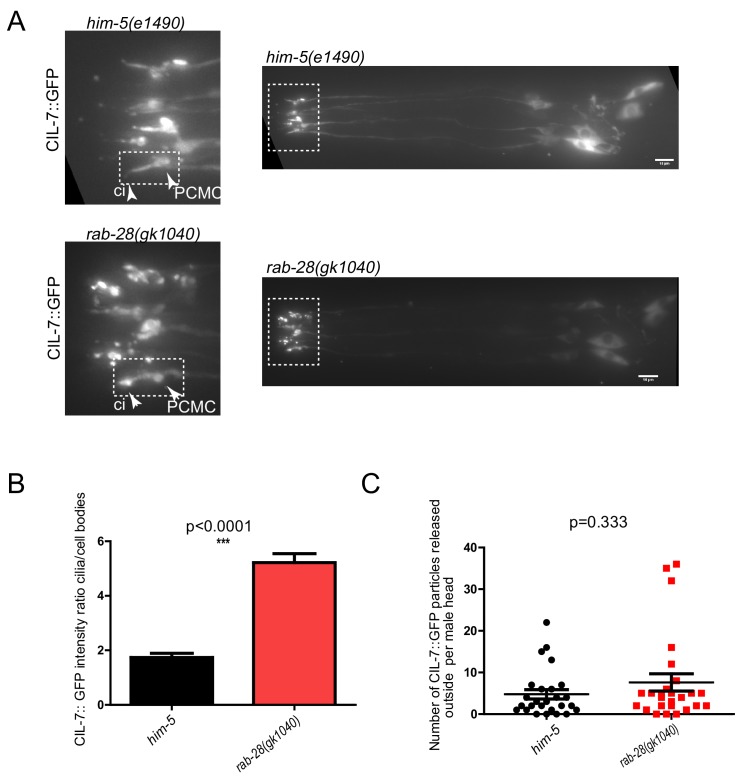
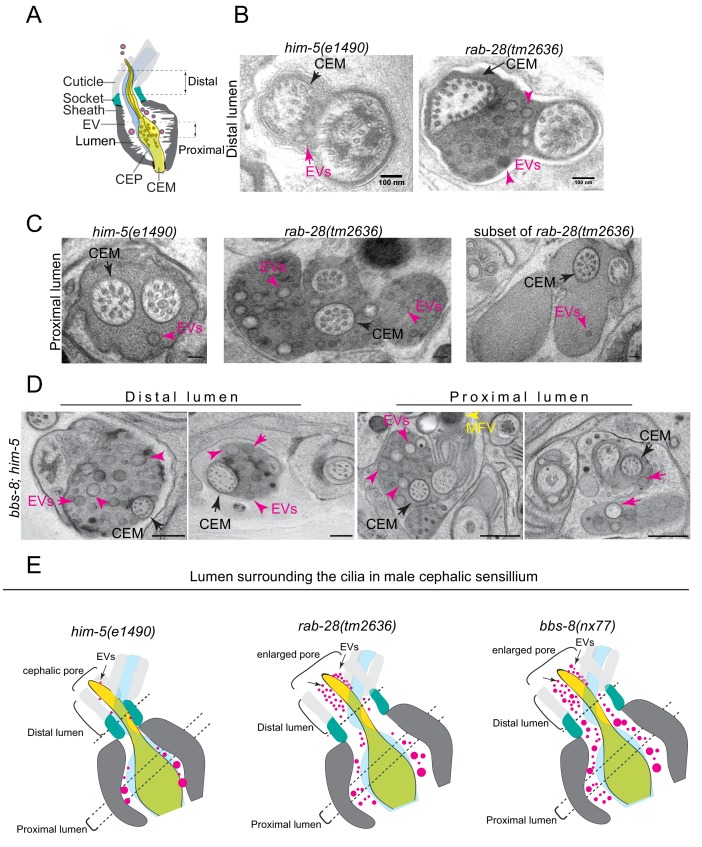
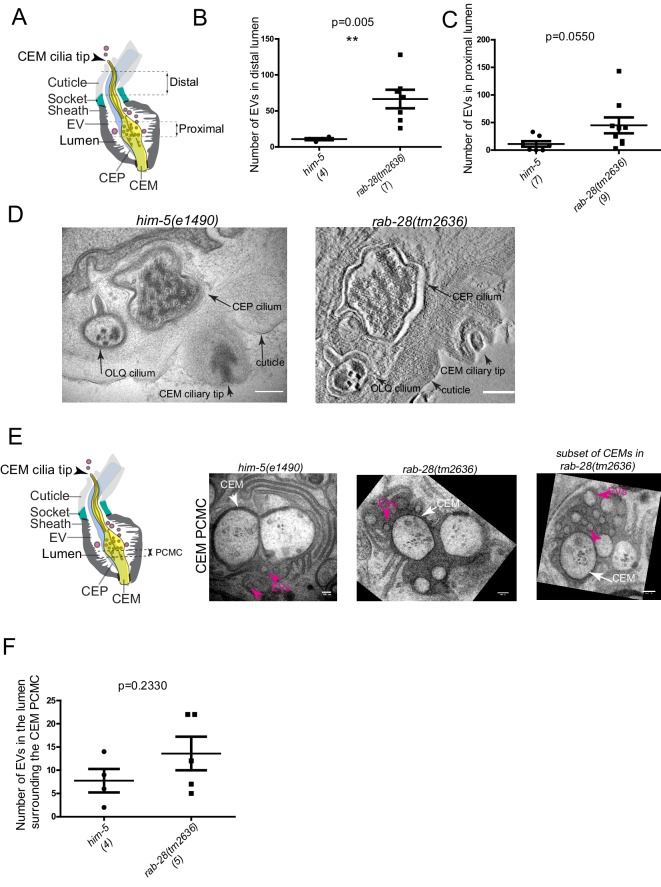
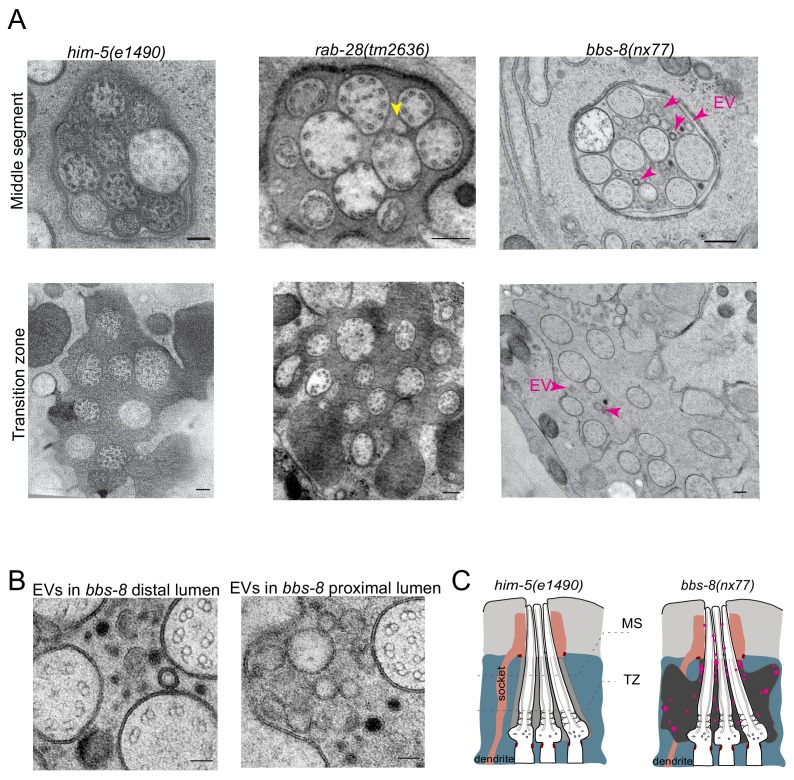
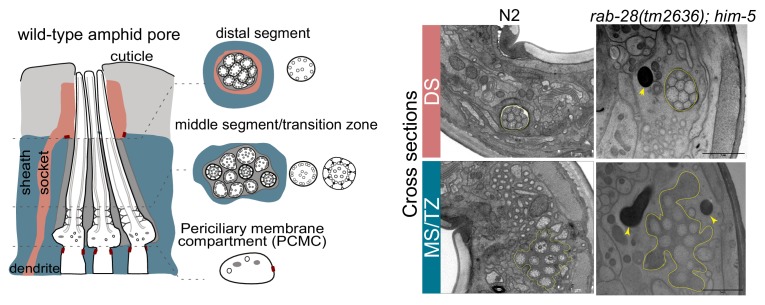
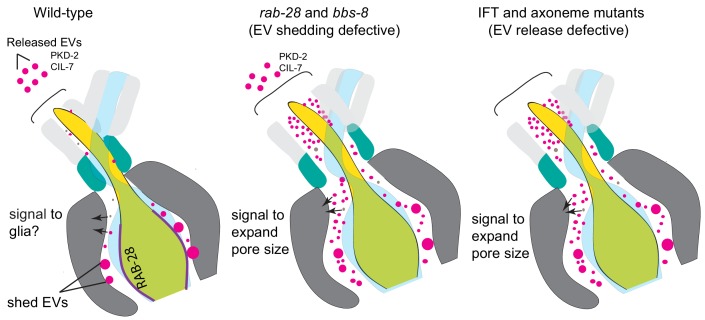
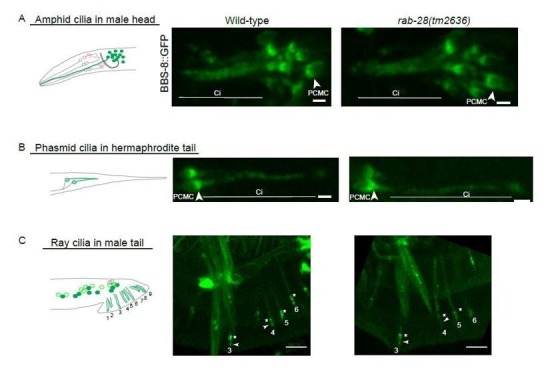
Cilia both receive and send information, the latter in the form of extracellular vesicles (EVs). EVs are nano-communication devices that influence cell, tissue, and organism behavior. Mechanisms driving ciliary EV biogenesis are almost entirely unknown. Here, we show that the ciliary G-protein Rab28, associated with human autosomal recessive cone-rod dystrophy, negatively regulates EV levels in the sensory organs of Caenorhabditis elegans in a cilia specific manner. Sequential targeting of lipidated Rab28 to periciliary and ciliary membranes is highly dependent on the BBSome and the prenyl-binding protein phosphodiesterase 6 subunit delta (PDE6D), respectively, and BBSome loss causes excessive and ectopic EV production. We also find that EV defective mutants display abnormalities in sensory compartment morphogenesis. Together, these findings reveal that Rab28 and the BBSome are key in vivo regulators of EV production at the periciliary membrane and suggest that EVs may mediate signaling between cilia and glia to shape sensory organ compartments. Our data also suggest that defects in the biogenesis of cilia-related EVs may contribute to human ciliopathies.
Keywords: BBSome; C. elegans; PKD2; Rab28; cell biology; cilia; ciliopathy; extracellular vesicles.
© 2020, Akella et al.
Conflict of interest statement
JA, SC, KN, ST, AM, MS, FR, BK, DH, MB, OB No competing interests declared
Figures
References
-
- Ansley SJ, Badano JL, Blacque OE, Hill J, Hoskins BE, Leitch CC, Kim JC, Ross AJ, Eichers ER, Teslovich TM, Mah AK, Johnsen RC, Cavender JC, Lewis RA, Leroux MR, Beales PL, Katsanis N. Basal body dysfunction is a likely cause of pleiotropic Bardet-Biedl syndrome. Nature. 2003;425:628–633. doi: 10.1038/nature02030. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
