

Complex Evolution of Insect Insulin Receptors and Homologous Decoy Receptors, and Functional Significance of Their Multiplicity
- PMID: 32101294
- PMCID: PMC7253209
- DOI: 10.1093/molbev/msaa048
Complex Evolution of Insect Insulin Receptors and Homologous Decoy Receptors, and Functional Significance of Their Multiplicity
Abstract
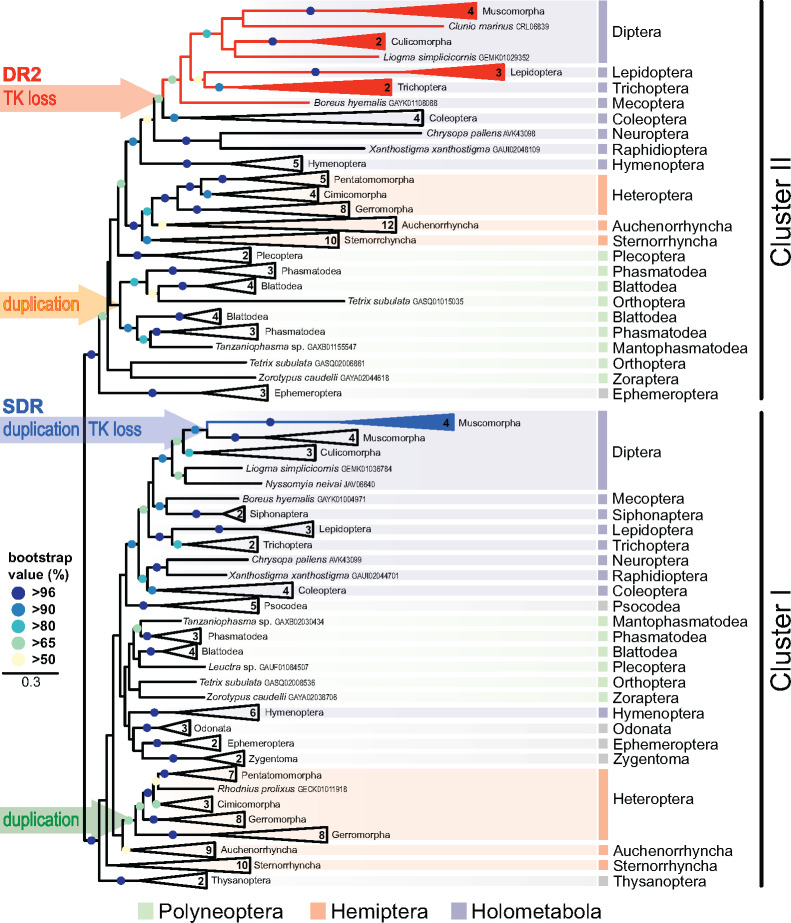
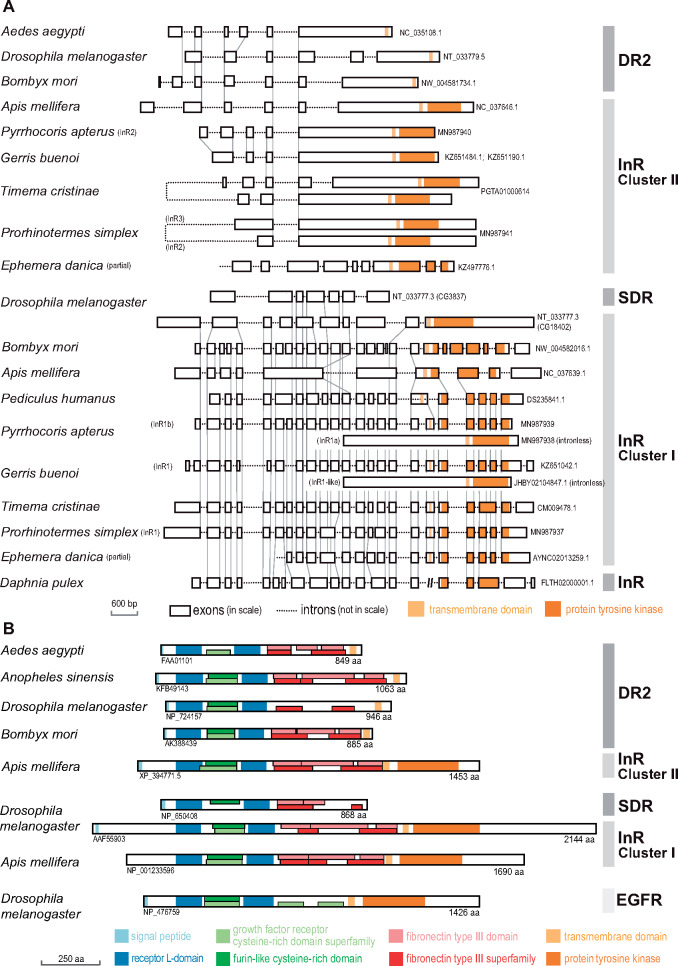
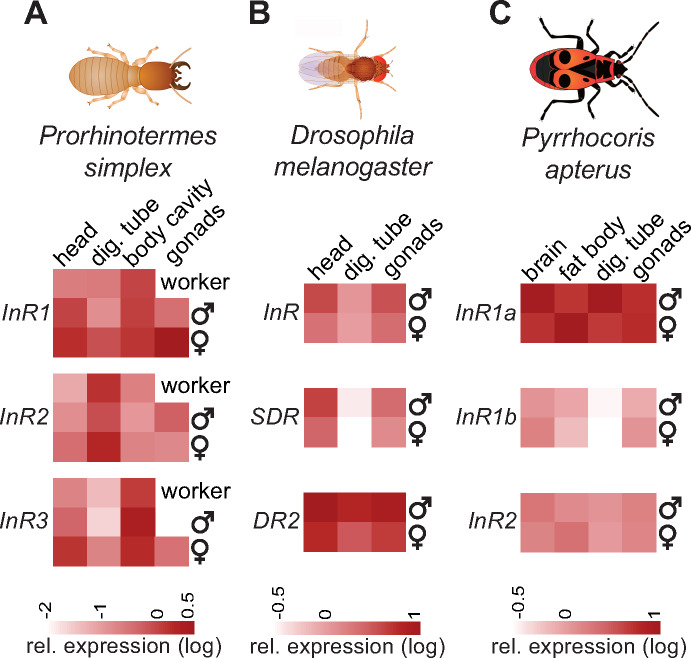
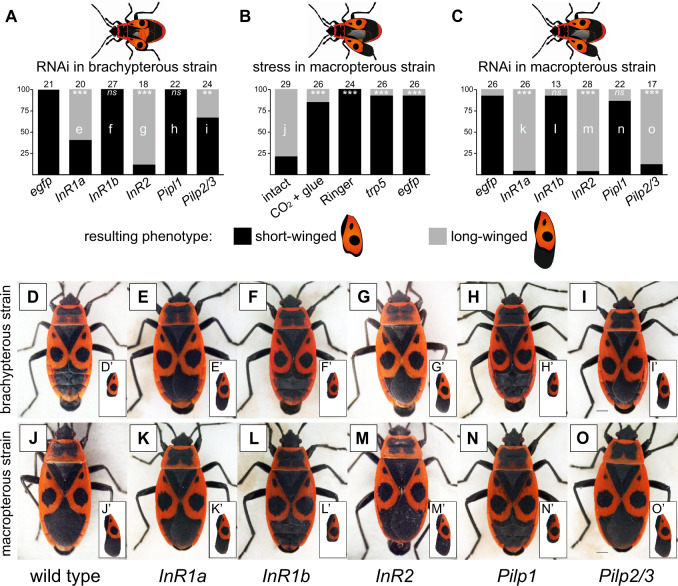
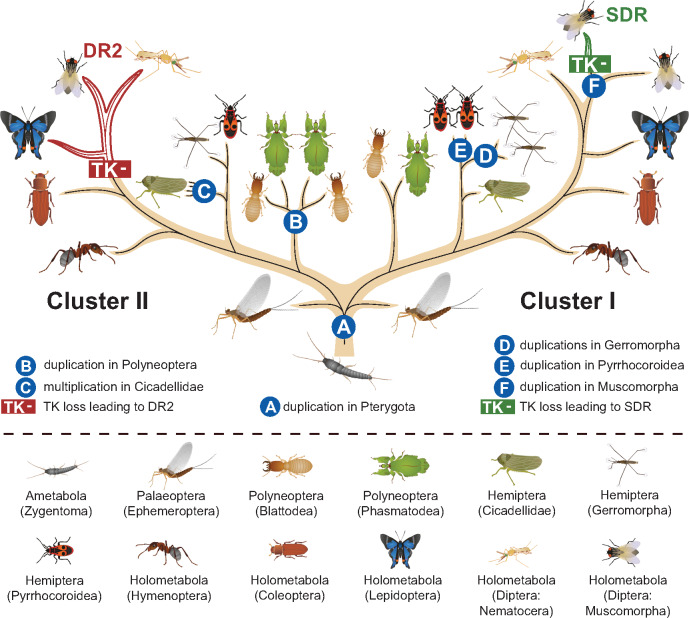
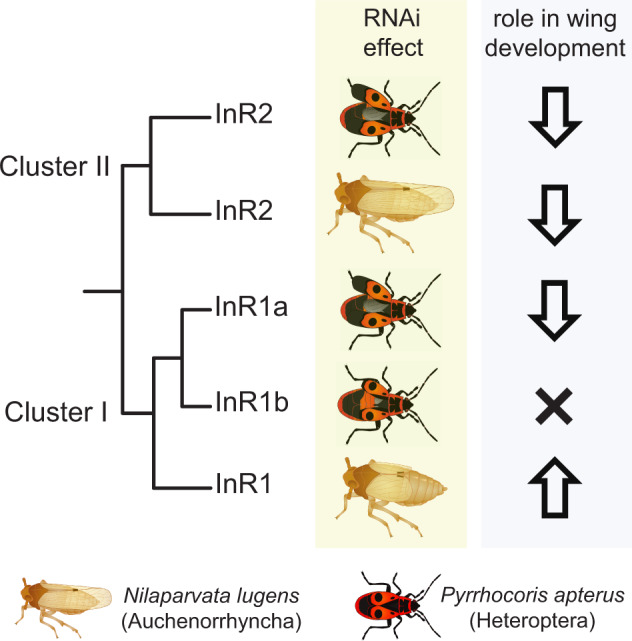
Evidence accumulates that the functional plasticity of insulin and insulin-like growth factor signaling in insects could spring, among others, from the multiplicity of insulin receptors (InRs). Their multiple variants may be implemented in the control of insect polyphenism, such as wing or caste polyphenism. Here, we present a comprehensive phylogenetic analysis of insect InR sequences in 118 species from 23 orders and investigate the role of three InRs identified in the linden bug, Pyrrhocoris apterus, in wing polymorphism control. We identified two gene clusters (Clusters I and II) resulting from an ancestral duplication in a late ancestor of winged insects, which remained conserved in most lineages, only in some of them being subject to further duplications or losses. One remarkable yet neglected feature of InR evolution is the loss of the tyrosine kinase catalytic domain, giving rise to decoys of InR in both clusters. Within the Cluster I, we confirmed the presence of the secreted decoy of insulin receptor in all studied Muscomorpha. More importantly, we described a new tyrosine kinase-less gene (DR2) in the Cluster II, conserved in apical Holometabola for ∼300 My. We differentially silenced the three P. apterus InRs and confirmed their participation in wing polymorphism control. We observed a pattern of Cluster I and Cluster II InRs impact on wing development, which differed from that postulated in planthoppers, suggesting an independent establishment of insulin/insulin-like growth factor signaling control over wing development, leading to idiosyncrasies in the co-option of multiple InRs in polyphenism control in different taxa.
Keywords: decoy of insulin receptor; gene structure; insects; insulin receptor; insulin signaling; wing polyphenism.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Abrisqueta M, Suren-Castillo S, Maestro JL.. 2014. Insulin receptor-mediated nutritional signalling regulates juvenile hormone biosynthesis and vitellogenin production in the German cockroach. Insect Biochem Mol Biol. 49:14–23. - PubMed
-
- Badisco L, Claeys I, Van Hiel M, Clynen E, Huybrechts J, Vandersmissen T, Van Soest S, Vanden Bosch L, Simonet G, Vanden Broeck J.. 2008. Purification and characterization of an insulin-related peptide in the desert locust, Schistocerca gregaria: immunolocalization, cDNA cloning, transcript profiling and interaction with neuroparsin. J Mol Endocrinol. 40(3):137–150. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
