

Establishment and optimization of a hemp (Cannabis sativa L.) agroinfiltration system for gene expression and silencing studies
- PMID: 32103049
- PMCID: PMC7044430
- DOI: 10.1038/s41598-020-60323-9
Establishment and optimization of a hemp (Cannabis sativa L.) agroinfiltration system for gene expression and silencing studies
Abstract
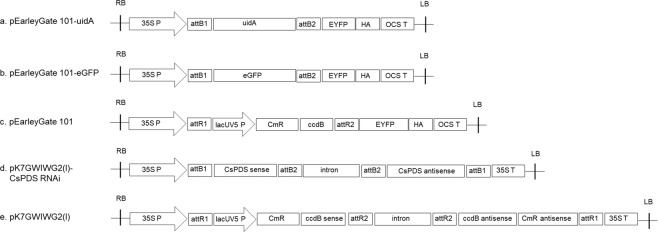
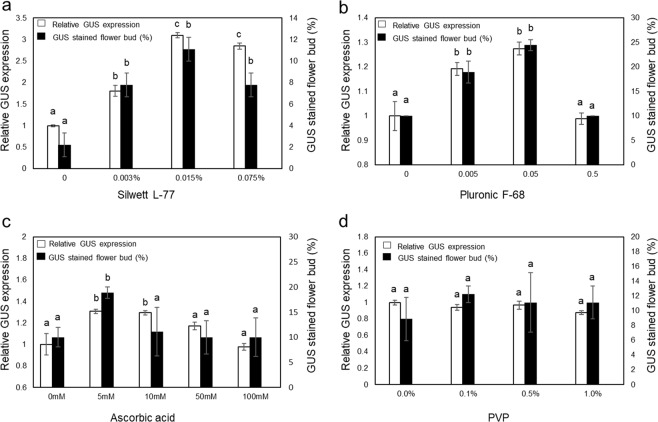
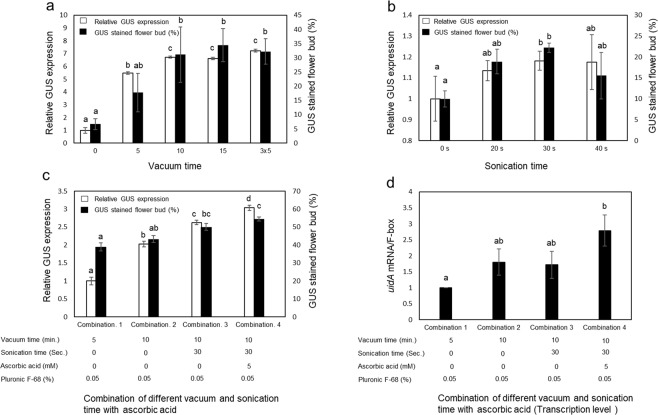
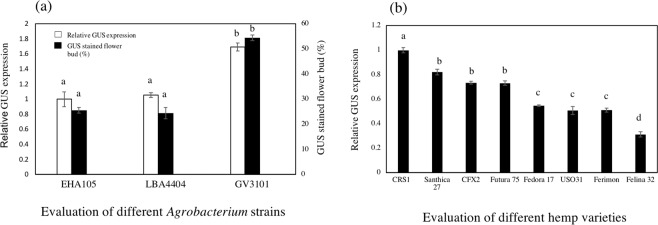
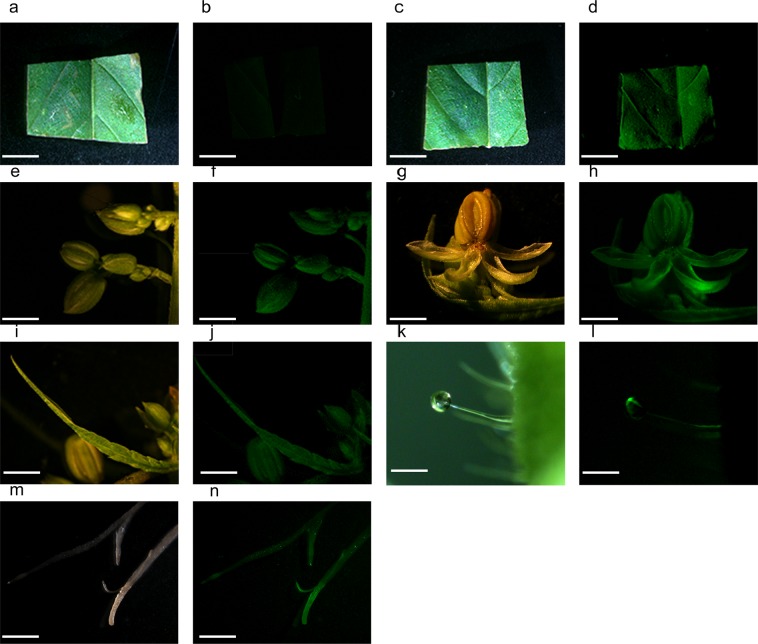
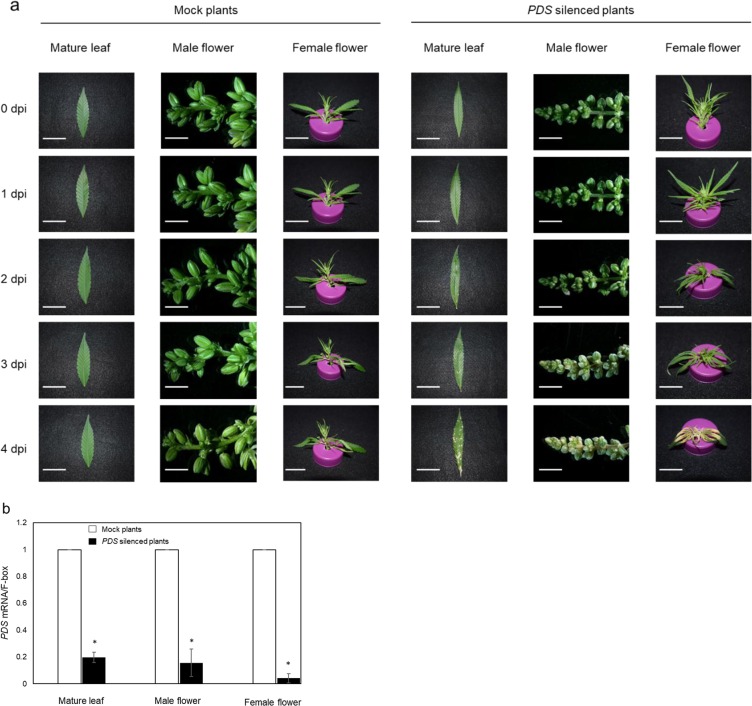
Industrial hemp (Cannabis sativa L.) is a high-yielding annual crop primarily grown for fiber, seeds, and oil. Due to the phytochemical composition of hemp, there has been an increased interest in the market for nutraceuticals and dietary supplements for human health. Recent omics analysis has led to the elucidation of hemp candidate genes involved in the syntheses of specialized metabolites. However, a detailed study of these genes has not been undertaken due to the lack of a stable transformation system. We report for the first time an agroinfiltration system in hemp utilizing vacuum infiltration, which is an alternative method to stable transformation. A combination of 0.015% Silwett L-77, 5 mM ascorbic acid, and thirty second sonication followed by a 10-minute vacuum treatment resulted in the highest β-glucuronidase expression in the leaf, male and female flowers, stem, and root tissues. The phytoene desaturase gene was silenced with a transient hairpin RNA expression, resulting in an albino phenotype in the leaves and the male and female flowers. This agroinfiltration system would be useful for overexpression and silencing studies of target genes to regulate the yield of specialized metabolites in hemp.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Chandra, S., Lata, H., Khan, I., & ElSohly, M. A. Cannabis Sativa L.: Botany and Horticulture in Cannabis sativa L.: Botany and Biotechnology. Cham. Switzerland: Springer. 79–100 (2017).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
