

Global chemical effects of the microbiome include new bile-acid conjugations
- PMID: 32103176
- PMCID: PMC7252668
- DOI: 10.1038/s41586-020-2047-9
Global chemical effects of the microbiome include new bile-acid conjugations
Abstract
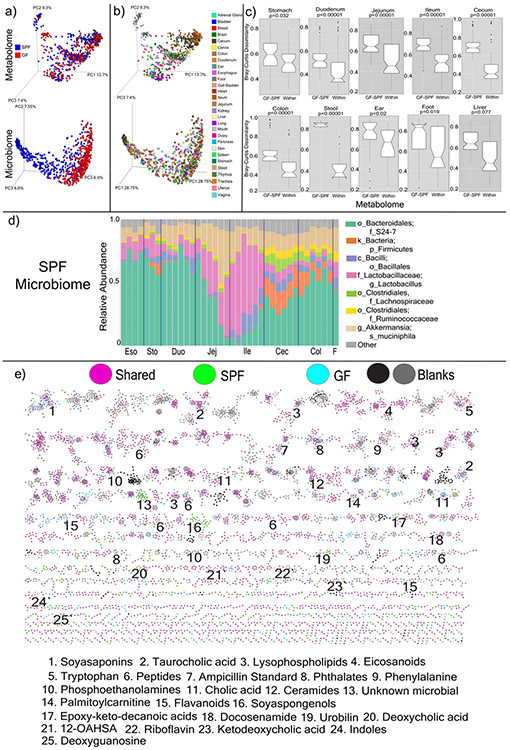
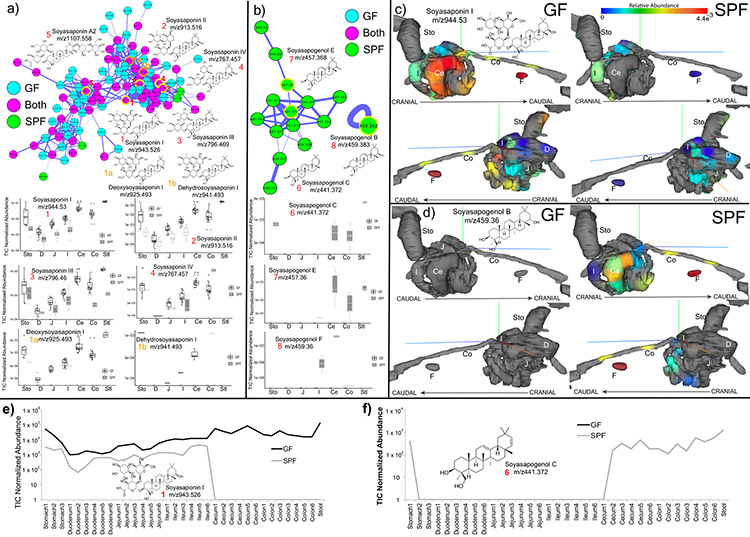
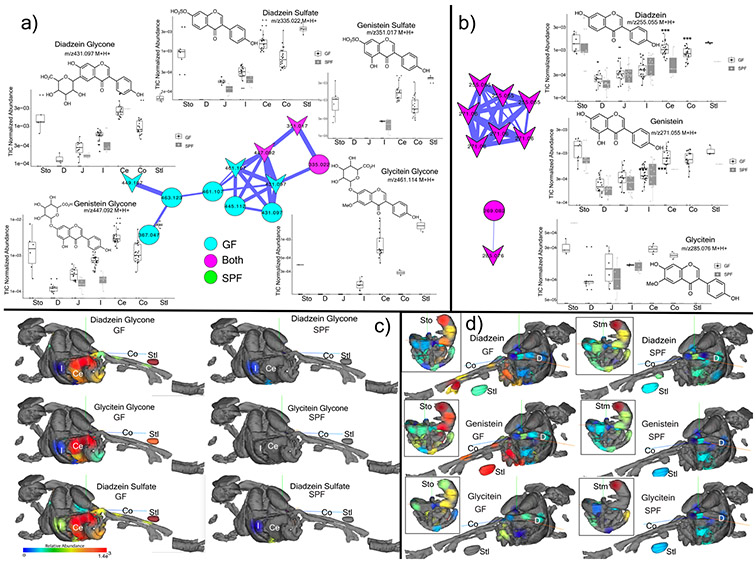
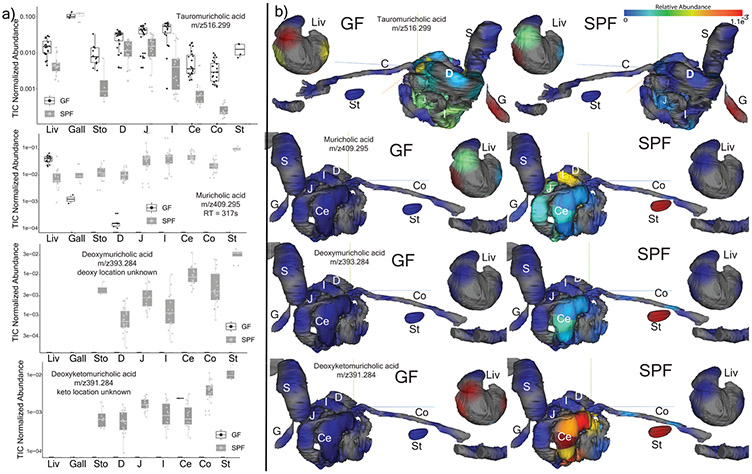
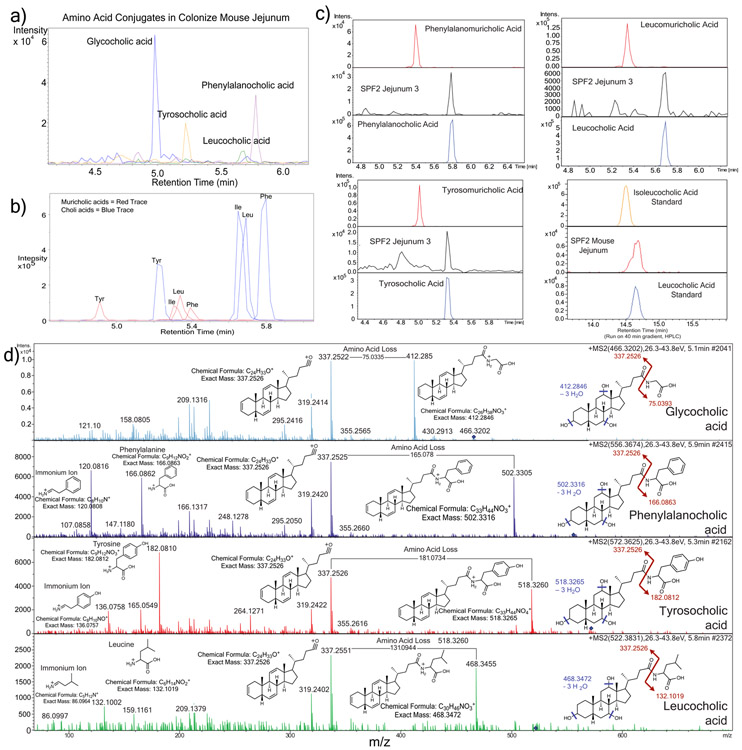
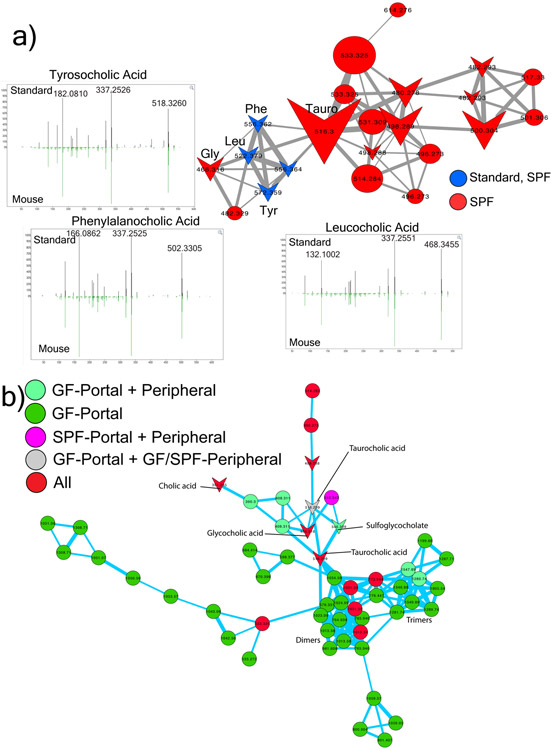
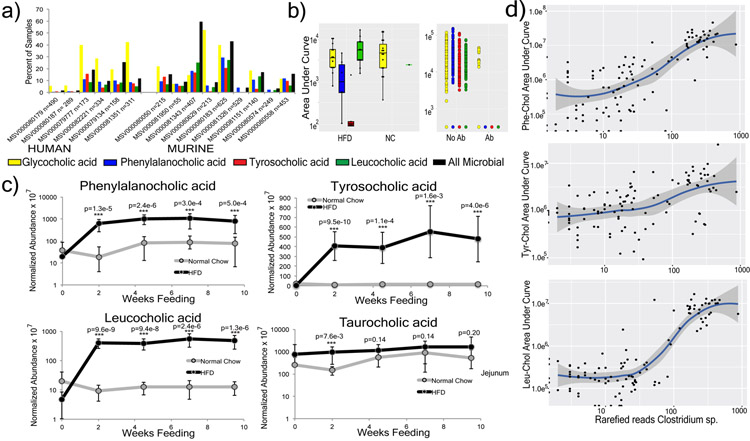
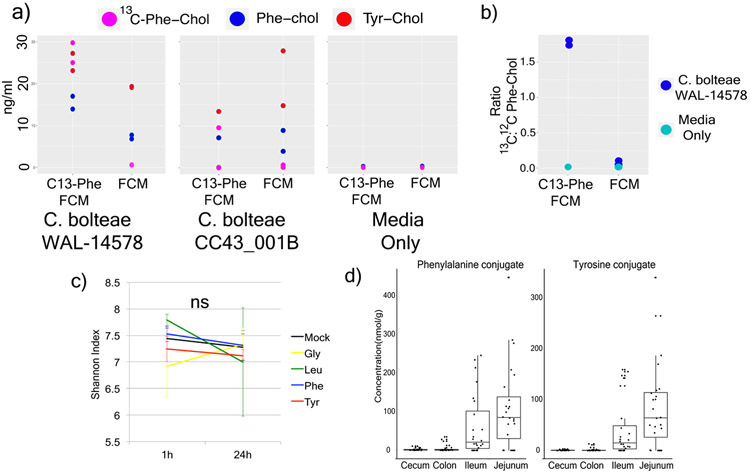
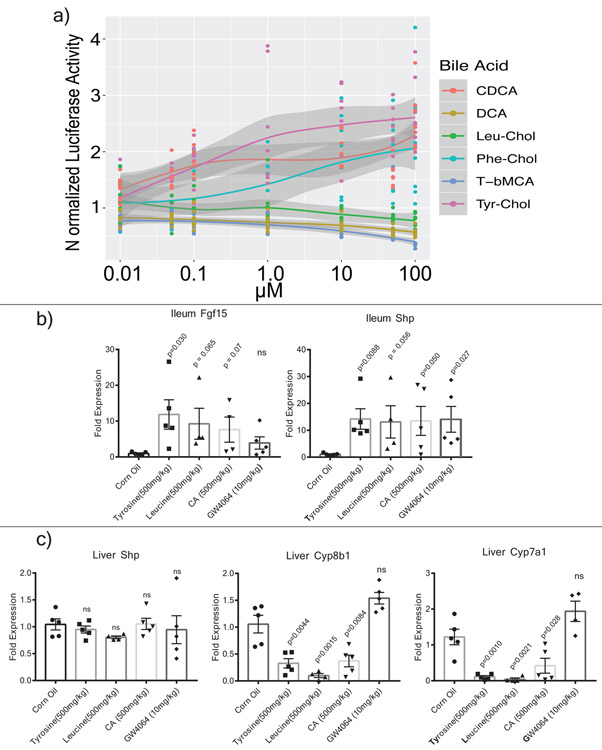
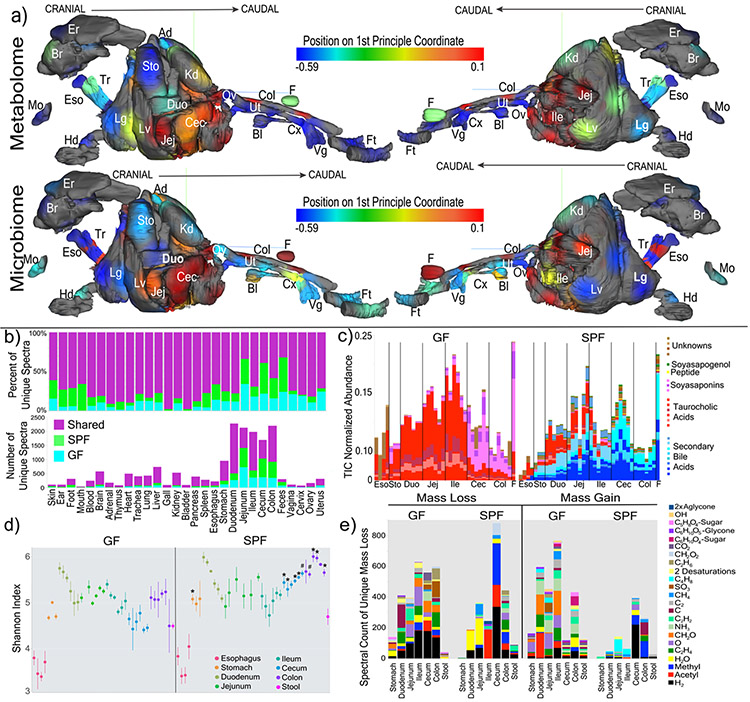
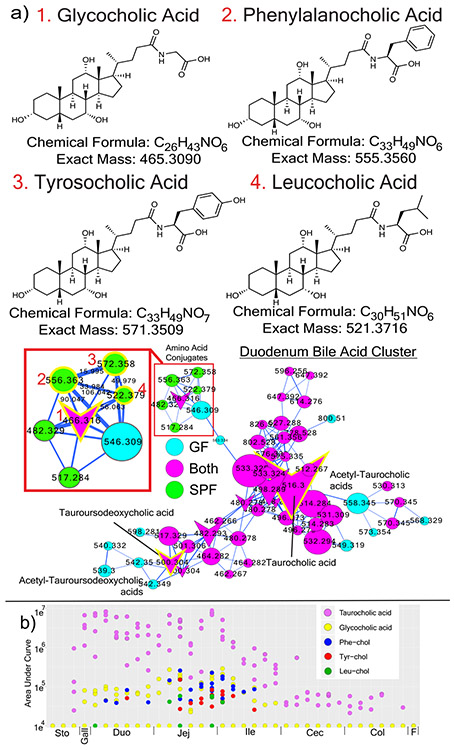
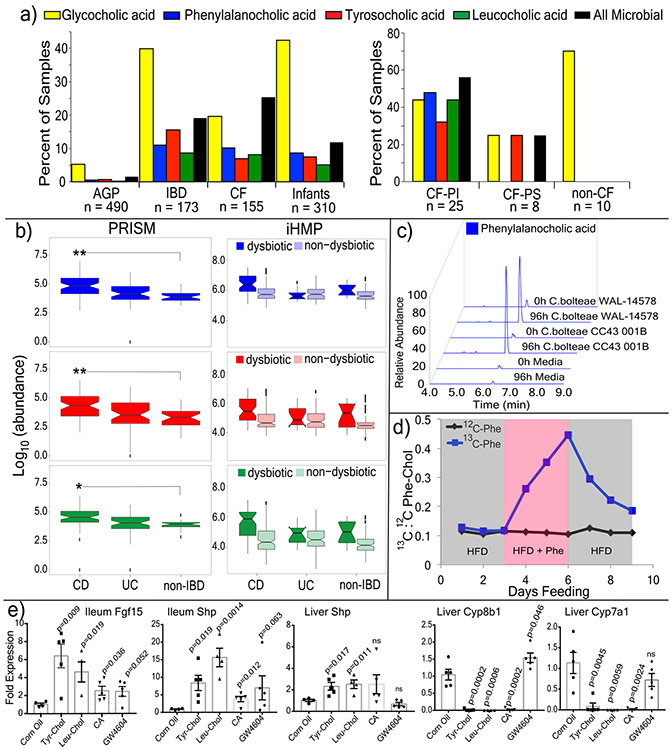
A mosaic of cross-phylum chemical interactions occurs between all metazoans and their microbiomes. A number of molecular families that are known to be produced by the microbiome have a marked effect on the balance between health and disease1-9. Considering the diversity of the human microbiome (which numbers over 40,000 operational taxonomic units10), the effect of the microbiome on the chemistry of an entire animal remains underexplored. Here we use mass spectrometry informatics and data visualization approaches11-13 to provide an assessment of the effects of the microbiome on the chemistry of an entire mammal by comparing metabolomics data from germ-free and specific-pathogen-free mice. We found that the microbiota affects the chemistry of all organs. This included the amino acid conjugations of host bile acids that were used to produce phenylalanocholic acid, tyrosocholic acid and leucocholic acid, which have not previously been characterized despite extensive research on bile-acid chemistry14. These bile-acid conjugates were also found in humans, and were enriched in patients with inflammatory bowel disease or cystic fibrosis. These compounds agonized the farnesoid X receptor in vitro, and mice gavaged with the compounds showed reduced expression of bile-acid synthesis genes in vivo. Further studies are required to confirm whether these compounds have a physiological role in the host, and whether they contribute to gut diseases that are associated with microbiome dysbiosis.
Figures
Comment in
-
Gut Microbes Take It to the Next Level? First Insights Into Farnesoid X Receptor Agonists of Microbial Origin.Hepatology. 2020 Oct;72(4):1483-1485. doi: 10.1002/hep.31455. Hepatology. 2020. PMID: 32648608 Free PMC article. No abstract available.
-
Microbially conjugated bile salts found in human bile activate the bile salt receptors TGR5 and FXR.Hepatol Commun. 2024 Mar 22;8(4):e0383. doi: 10.1097/HC9.0000000000000383. eCollection 2024 Apr 1. Hepatol Commun. 2024. PMID: 38517202 Free PMC article.
References
-
- Gilbert JA et al. Microbiome-wide association studies link dynamic microbial consortia to disease. Nature 535, (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL088093/HL/NHLBI NIH HHS/United States
- GMS10RR029121/NH/NIH HHS/United States
- R24DK110499/NH/NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- 1 DP1 AT010885/NH/NIH HHS/United States
- 1R01HL116235/NH/NIH HHS/United States
- KL2 TR001444/TR/NCATS NIH HHS/United States
- R37 DK057978/DK/NIDDK NIH HHS/United States
- T32 DK007494/DK/NIDDK NIH HHS/United States
- R03 CA211211/CA/NCI NIH HHS/United States
- R24 DK110499/DK/NIDDK NIH HHS/United States
- P30 DK040561/DK/NIDDK NIH HHS/United States
- R01 HL105278/HL/NHLBI NIH HHS/United States
- R01 HD084163/HD/NICHD NIH HHS/United States
- R01 GM107550/GM/NIGMS NIH HHS/United States
- S10 RR029121/RR/NCRR NIH HHS/United States
- CA014195/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- U01 AI124316/AI/NIAID NIH HHS/United States
- P42 ES010337/ES/NIEHS NIH HHS/United States
- R01 AI126277/AI/NIAID NIH HHS/United States
- DP1 AT010885/AT/NCCIH NIH HHS/United States
- 1R03CA211211-01/NH/NIH HHS/United States
- P30 DK120515/DK/NIDDK NIH HHS/United States
- U54 DE023798/DE/NIDCR NIH HHS/United States
- R01 HL116235/HL/NHLBI NIH HHS/United States
- T15 LM011271/LM/NLM NIH HHS/United States
- U54DE023798/NH/NIH HHS/United States
- 5U01AI124316-03/NH/NIH HHS/United States
- R01 DK057978/DK/NIDDK NIH HHS/United States
- P30 CA014195/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
