

IKKε phosphorylates kindlin-2 to induce invadopodia formation and promote colorectal cancer metastasis
- PMID: 32104508
- PMCID: PMC7019159
- DOI: 10.7150/thno.40397
IKKε phosphorylates kindlin-2 to induce invadopodia formation and promote colorectal cancer metastasis
Abstract
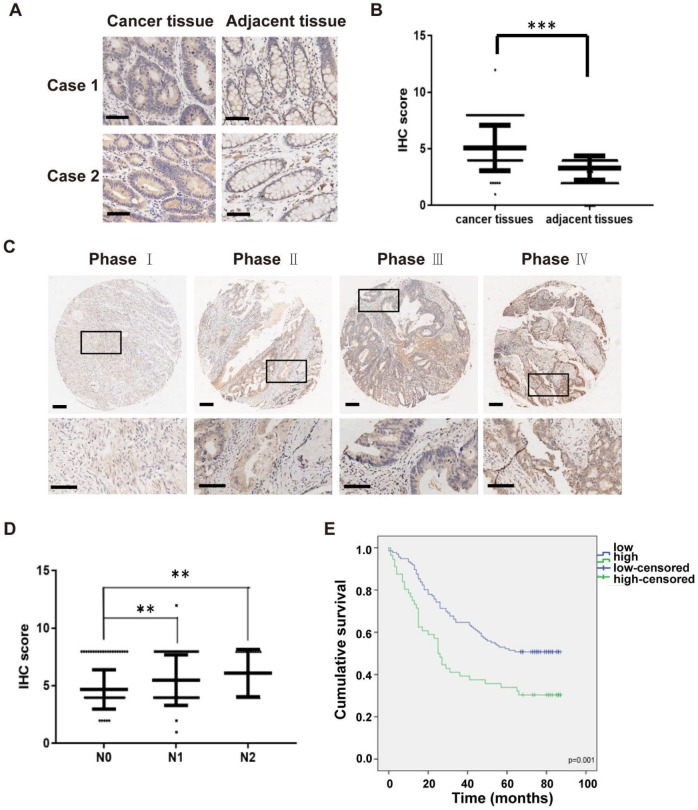
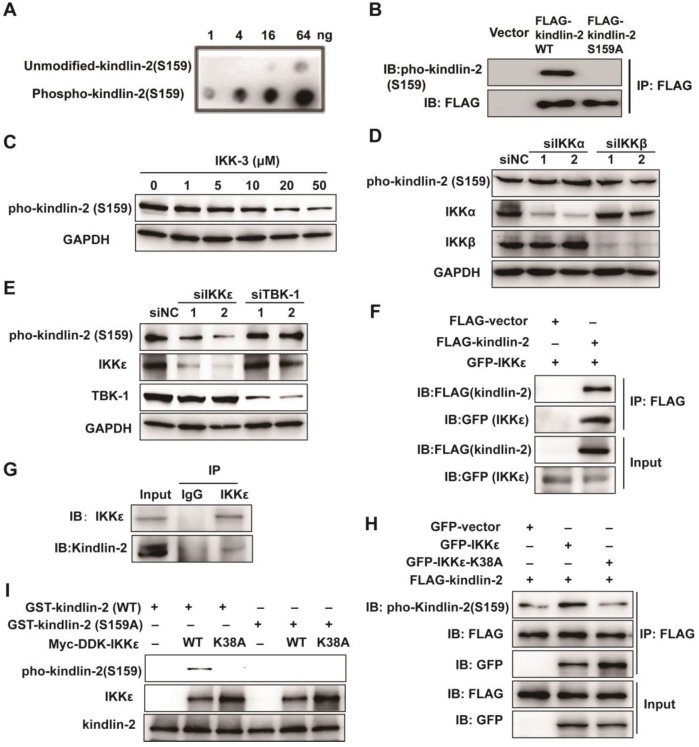
Invadopodia formation is a key driver of cancer metastasis. The noncanonical IkB-related kinase IKKε has been implicated in cancer metastasis, but its roles in invadopodia formation and colorectal cancer (CRC) metastasis are unclear. Methods: Immunofluorescence, gelatin-degradation assay, wound healing assay and transwell invasion assay were used to determine the influence of IKKε over-expression, knockdown and pharmacological inhibition on invadopodia formation and the migratory and invasive capacity of CRC cells in vitro. Effects of IKKε knockdown or pharmacological inhibition on CRC metastasis were examined in mice. Immunohistochemistry staining was used to detect expression levels of IKKε in CRC patient tissues, and its association with prognosis in CRC patients was also analyzed. Immunoprecipitation, western blotting and in vitro kinase assay were constructed to investigate the molecular mechanisms. Results: IKKε co-localizes with F-actin and the invadopodia marker Tks5 at the gelatin-degrading sites of CRC cells. Genetic over-expression/knockdown or pharmacological inhibition of IKKε altered invadopodia formation and the migratory and invasive capacity of CRC cells in vitro. In vivo, knockdown or pharmacological inhibition of IKKε significantly suppressed metastasis of CRC cells in mice. IKKε knockdown also inhibited invadopodia formation in vivo. Clinical investigation of tumor specimens from 191 patients with CRC revealed that high IKKε expression correlates with metastasis and poor prognosis of CRC. Mechanistically, IKKε directly binds to and phosphorylates kindlin-2 at serine 159; this effect mediates the IKKε-induced invadopodia formation and promotion of CRC metastasis. Conclusions: We identify IKKε as a novel regulator of invadopodia formation and a unique mechanism by which IKKε promotes the metastasis of CRC. Our study suggests that IKKε is a potential target to suppress CRC metastasis.
Keywords: Colorectal cancer; IKKε; Invadopodia; Kindlin-2 phosphorylation; Metastasis.
© The author(s).
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures






Similar articles
-
Hyper activation of β-catenin signalling induced by IKKε inhibition thwarts colorectal cancer cell proliferation.Cell Prolif. 2017 Aug;50(4):e12350. doi: 10.1111/cpr.12350. Epub 2017 May 19. Cell Prolif. 2017. PMID: 28523736 Free PMC article.
-
Cortactin recruits FMNL2 to promote actin polymerization and endosome motility in invadopodia formation.Cancer Lett. 2018 Apr 10;419:245-256. doi: 10.1016/j.canlet.2018.01.023. Epub 2018 Jan 31. Cancer Lett. 2018. PMID: 29374558
-
Silencing of IKKε using siRNA inhibits proliferation and invasion of glioma cells in vitro and in vivo.Int J Oncol. 2012 Jul;41(1):169-78. doi: 10.3892/ijo.2012.1452. Epub 2012 Apr 26. Int J Oncol. 2012. PMID: 22552702
-
The involvement of mutant Rac1 in the formation of invadopodia in cultured melanoma cells.Exp Cell Res. 2016 Apr 10;343(1):82-88. doi: 10.1016/j.yexcr.2016.02.003. Epub 2016 Feb 10. Exp Cell Res. 2016. PMID: 26873115 Free PMC article. Review.
-
Invadopodia in cancer metastasis: dynamics, regulation, and targeted therapies.J Transl Med. 2025 May 16;23(1):548. doi: 10.1186/s12967-025-06526-y. J Transl Med. 2025. PMID: 40380267 Free PMC article. Review.
Cited by
-
Migfilin: Cell Adhesion Effect and Comorbidities.Onco Targets Ther. 2022 Apr 19;15:411-422. doi: 10.2147/OTT.S357355. eCollection 2022. Onco Targets Ther. 2022. PMID: 35469339 Free PMC article. Review.
-
Kindlin-2 enhances c-Myc translation through association with DDX3X to promote pancreatic ductal adenocarcinoma progression.Theranostics. 2023 Jul 31;13(13):4333-4355. doi: 10.7150/thno.85421. eCollection 2023. Theranostics. 2023. PMID: 37649609 Free PMC article.
-
Role of Post-Translational Modifications in Colorectal Cancer Metastasis.Cancers (Basel). 2024 Feb 3;16(3):652. doi: 10.3390/cancers16030652. Cancers (Basel). 2024. PMID: 38339403 Free PMC article. Review.
-
Phosphorylation of Kindlins and the Control of Integrin Function.Cells. 2021 Apr 7;10(4):825. doi: 10.3390/cells10040825. Cells. 2021. PMID: 33916922 Free PMC article. Review.
-
Targeting the JAK-STAT pathway in colorectal cancer: mechanisms, clinical implications, and therapeutic potential.Front Cell Dev Biol. 2024 Nov 26;12:1507621. doi: 10.3389/fcell.2024.1507621. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39659524 Free PMC article. Review.
References
-
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. - PubMed
-
- Yamaguchi H. Pathological roles of invadopodia in cancer invasion and metastasis. Eur J Cell Biol. 2012;91:902–7. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
