

The cell biology of the retinal pigment epithelium
- PMID: 32105772
- PMCID: PMC8941496
- DOI: 10.1016/j.preteyeres.2020.100846
The cell biology of the retinal pigment epithelium
Abstract
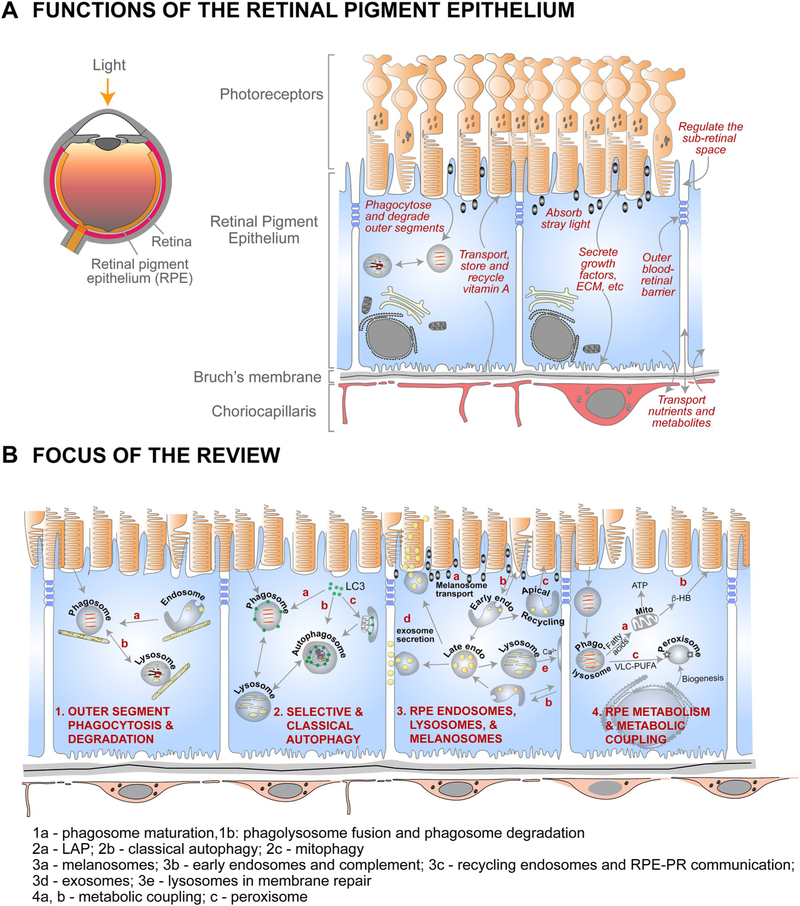
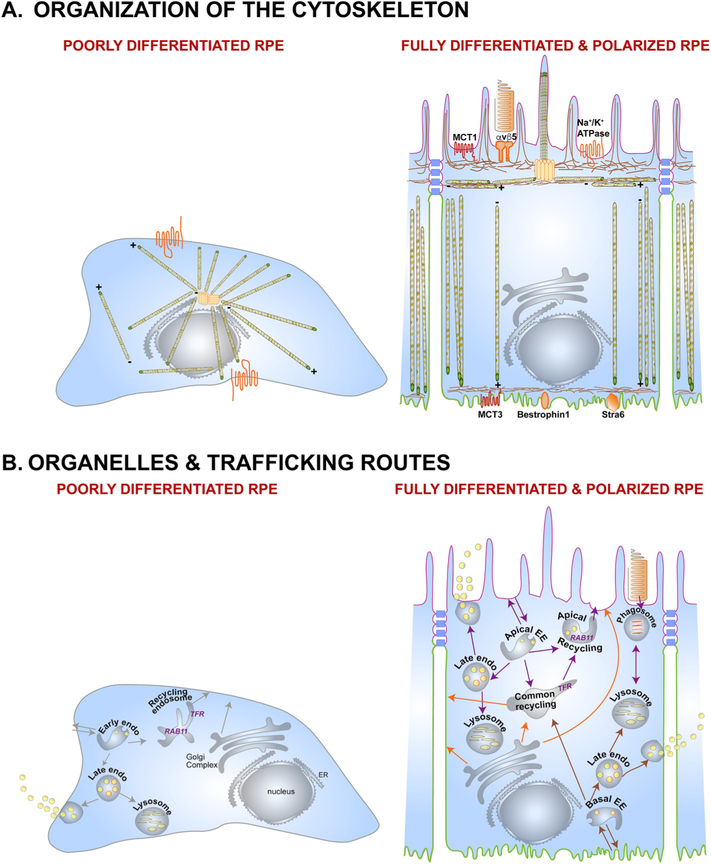
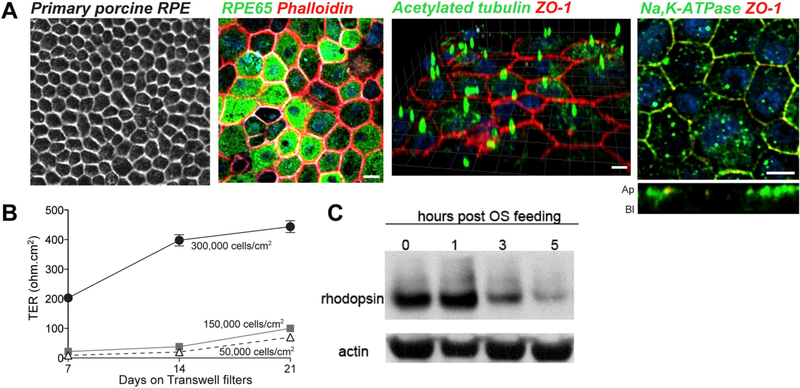
The retinal pigment epithelium (RPE), a monolayer of post-mitotic polarized epithelial cells, strategically situated between the photoreceptors and the choroid, is the primary caretaker of photoreceptor health and function. Dysfunction of the RPE underlies many inherited and acquired diseases that cause permanent blindness. Decades of research have yielded valuable insight into the cell biology of the RPE. In recent years, new technologies such as live-cell imaging have resulted in major advancement in our understanding of areas such as the daily phagocytosis and clearance of photoreceptor outer segment tips, autophagy, endolysosome function, and the metabolic interplay between the RPE and photoreceptors. In this review, we aim to integrate these studies with an emphasis on appropriate models and techniques to investigate RPE cell biology and metabolism, and discuss how RPE cell biology informs our understanding of retinal disease.
Keywords: Autophagy; Endosomes; Lysosomes; Metabolism; Organelles; Phagocytosis; Phagosome maturation.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

References
-
- Ahmado A, Carr AJ, Vugler AA, Semo M, Gias C, Lawrence JM, Chen LL, Chen FK, Turowski P, da Cruz L, Coffey PJ, 2011. Induction of differentiation by pyruvate and DMEM in the human retinal pigment epithelium cell line ARPE-19. Invest. Ophthalmol. Vis. Sci. 52, 7148–7159. 10.1167/iovs.10-6374. - DOI - PubMed
Publication types
Grants and funding
- R01 EY023299/EY/NEI NIH HHS/United States
- R21 EY029826/EY/NEI NIH HHS/United States
- R01 EY010420/EY/NEI NIH HHS/United States
- P30 EY000331/EY/NEI NIH HHS/United States
- R01 EY026525/EY/NEI NIH HHS/United States
- R21 EY032743/EY/NEI NIH HHS/United States
- R01 EY012042/EY/NEI NIH HHS/United States
- P30 EY001583/EY/NEI NIH HHS/United States
- R01 EY027442/EY/NEI NIH HHS/United States
- R01 EY030668/EY/NEI NIH HHS/United States
- P30 EY002162/EY/NEI NIH HHS/United States
- R21 EY031109/EY/NEI NIH HHS/United States
- R01 EY013408/EY/NEI NIH HHS/United States
