

The effect of sediment grain properties and porewater flow on microbial abundance and respiration in permeable sediments
- PMID: 32107429
- PMCID: PMC7046789
- DOI: 10.1038/s41598-020-60557-7
The effect of sediment grain properties and porewater flow on microbial abundance and respiration in permeable sediments
Abstract
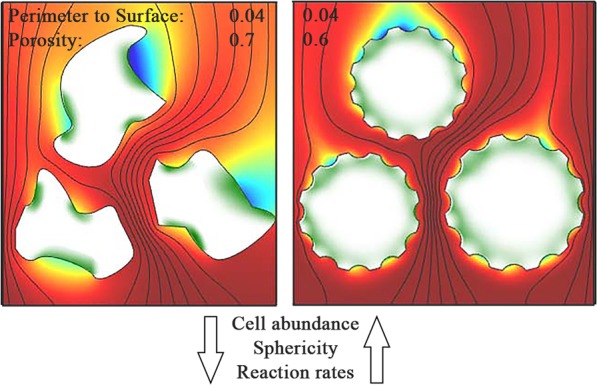
Sandy sediments cover 50-60% of the continental shelves and are highly efficient bioreactors in which organic carbon is remineralized and inorganic nitrogen is reduced to N2. As such they seem to play an important role, buffering the open ocean from anthropogenic nitrogen inputs and likely remineralizing the vast amounts of organic matter formed in the highly productive surface waters. To date however, little is known about the interrelation between porewater transport, grain properties and microbial colonization and the consequences for remineralization rates in sandy sediments. To constrain the effect of theses factors on remineralization in silicate sands, we incubated North Sea sediments in flow-through reactors after separating into five different grain size fractions. Bulk sediment and sediment grain properties were measured along with microbial colonization and cell abundances, oxygen consumption and denitrification rates. Volumetric oxygen consumption ranged from 14 to 77 µmol O2 l-1 h-1 while nitrogen-loss via denitrification was between 3.7 and 8.4 µmol N l-1 h-1. Oxygen consumption and denitrification rates were linearly correlated to the microbial cell abundances, which ranged from 2.9 to 5.4·108 cells cm-3. We found, that cell abundance and consumption rates in sandy sediments are influenced (i) by the surface area available for microbial colonization and (ii) by the exposure of these surfaces to the solute-supplying porewater flow. While protective structures such as cracks and depressions promote microbial colonization, the oxygen demand is only met by good ventilation of these structures, which is supported by a high sphericity of the grains. Based on our results, spherical sand grains with small depressions, i.e. golf ball like structures, provide the optimal supporting mineral structure for microorganisms on continental shelves.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Boudreau BP, et al. Permeable marine sediments: overturning an old paradigm. EOS, Transactions American Geophysical Union. 2001;82:133–136. doi: 10.1029/EO082i011p00133-01. - DOI
-
- Santos IR, Eyre BD, Huettel M. The driving forces of porewater and groundwater flow in permeable coastal sediments: A review. Estuarine, Coastal and Shelf Science. 2012;98:1–15. doi: 10.1016/j.ecss.2011.10.024. - DOI
-
- Ahmerkamp Soeren, Winter Christian, Krämer Knut, Beer Dirk de, Janssen Felix, Friedrich Jana, Kuypers Marcel M. M., Holtappels Moritz. Regulation of benthic oxygen fluxes in permeable sediments of the coastal ocean. Limnology and Oceanography. 2017;62(5):1935–1954. doi: 10.1002/lno.10544. - DOI
-
- Marchant HK, et al. Coupled nitrification–denitrification leads to extensive N loss in subtidal permeable sediments. Limnology and Oceanography. 2016;61:1033–1048. doi: 10.1002/lno.10271. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
