

Neuroestrogen synthesis modifies neural representations of learned song without altering vocal imitation in developing songbirds
- PMID: 32108169
- PMCID: PMC7046723
- DOI: 10.1038/s41598-020-60329-3
Neuroestrogen synthesis modifies neural representations of learned song without altering vocal imitation in developing songbirds
Abstract
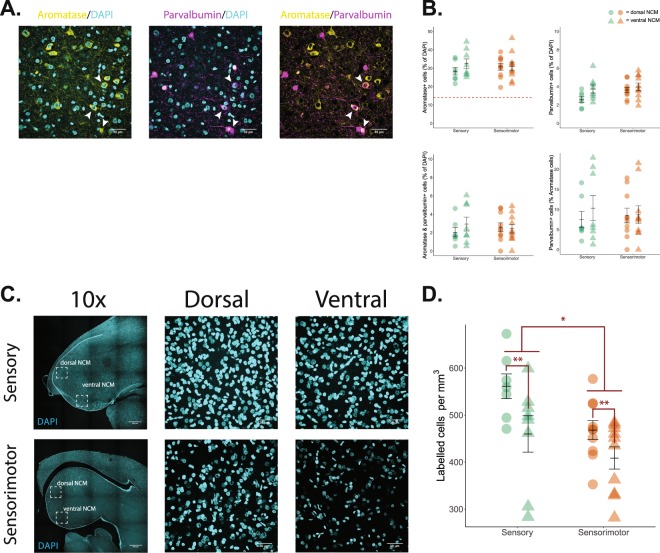
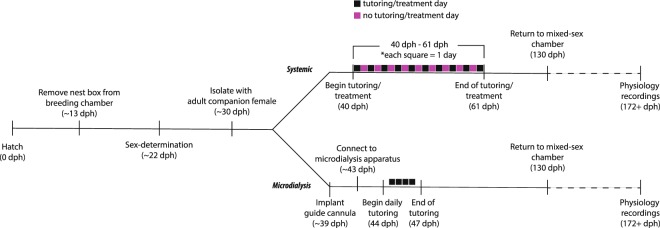
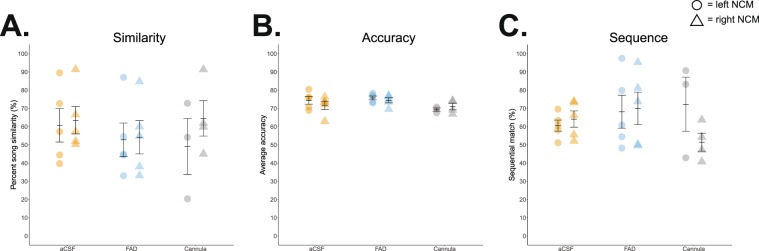
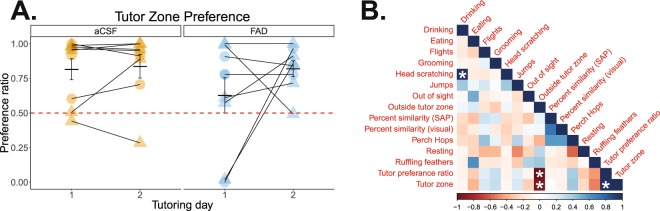
Birdsong learning, like human speech, depends on the early memorization of auditory models, yet how initial auditory experiences are formed and consolidated is unclear. In songbirds, a putative cortical locus is the caudomedial nidopallium (NCM), and one mechanism to facilitate auditory consolidation is 17β-estradiol (E2), which is associated with human speech-language development, and is abundant in both NCM and human temporal cortex. Circulating and NCM E2 levels are dynamic during learning, suggesting E2's involvement in encoding recent auditory experiences. Therefore, we tested this hypothesis in juvenile male songbirds using a comprehensive assessment of neuroanatomy, behavior, and neurophysiology. First, we found that brain aromatase expression, and thus the capacity to synthesize neuroestrogens, remains high in the auditory cortex throughout development. Further, while systemic estrogen synthesis blockade suppressed juvenile song production, neither systemic nor unilateral E2 synthesis inhibition in NCM disrupted eventual song imitation. Surprisingly, early life neuroestrogen synthesis blockade in NCM enhanced the neural representations of both the birds' own song and the tutor song in NCM and a downstream sensorimotor region, HVC, respectively. Taken together, these findings indicate that E2 plays a multifaceted role during development, and that, contrary to prediction, tutor song memorization is unimpaired by unilateral estrogen synthesis blockade in the auditory cortex.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Auditory learning in an operant task with social reinforcement is dependent on neuroestrogen synthesis in the male songbird auditory cortex.Horm Behav. 2020 May;121:104713. doi: 10.1016/j.yhbeh.2020.104713. Epub 2020 Feb 19. Horm Behav. 2020. PMID: 32057821 Free PMC article.
-
Neuroestrogen signaling in the songbird auditory cortex propagates into a sensorimotor network via an 'interface' nucleus.Neuroscience. 2015 Jan 22;284:522-535. doi: 10.1016/j.neuroscience.2014.10.023. Epub 2014 Oct 19. Neuroscience. 2015. PMID: 25453773 Free PMC article.
-
Sensory Coding and Sensitivity to Local Estrogens Shift during Critical Period Milestones in the Auditory Cortex of Male Songbirds.eNeuro. 2017 Dec 12;4(6):ENEURO.0317-17.2017. doi: 10.1523/ENEURO.0317-17.2017. eCollection 2017 Nov-Dec. eNeuro. 2017. PMID: 29255797 Free PMC article.
-
Birdsong memory and the brain: in search of the template.Neurosci Biobehav Rev. 2015 Mar;50:41-55. doi: 10.1016/j.neubiorev.2014.11.019. Epub 2014 Nov 29. Neurosci Biobehav Rev. 2015. PMID: 25459663 Review.
-
Brains for birds and babies: Neural parallels between birdsong and speech acquisition.Neurosci Biobehav Rev. 2017 Oct;81(Pt B):225-237. doi: 10.1016/j.neubiorev.2016.12.035. Epub 2017 Jan 10. Neurosci Biobehav Rev. 2017. PMID: 28087242 Review.
Cited by
-
Adult-like neural representation of species-specific songs in the auditory forebrain of zebra finch nestlings.Dev Neurobiol. 2021 Mar;81(2):123-138. doi: 10.1002/dneu.22802. Epub 2021 Jan 29. Dev Neurobiol. 2021. PMID: 33369121 Free PMC article.
-
Estrogen and sex-dependent loss of the vocal learning system in female zebra finches.Horm Behav. 2021 Mar;129:104911. doi: 10.1016/j.yhbeh.2020.104911. Epub 2021 Jan 23. Horm Behav. 2021. PMID: 33422557 Free PMC article.
-
Non-sensory Influences on Auditory Learning and Plasticity.J Assoc Res Otolaryngol. 2022 Apr;23(2):151-166. doi: 10.1007/s10162-022-00837-3. Epub 2022 Mar 2. J Assoc Res Otolaryngol. 2022. PMID: 35235100 Free PMC article. Review.
-
Dehydroepiandrosterone (DHEA) increases undirected singing behavior and alters dopaminergic regulation of undirected song in non-breeding male European starlings (Sturnus vulgaris).Front Endocrinol (Lausanne). 2023 May 10;14:1153085. doi: 10.3389/fendo.2023.1153085. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37234810 Free PMC article.
-
Social and auditory experience shapes forebrain responsiveness in zebra finches before the sensitive period of vocal learning.J Exp Biol. 2024 Nov 1;227(21):jeb247956. doi: 10.1242/jeb.247956. Epub 2024 Oct 25. J Exp Biol. 2024. PMID: 39263850 Free PMC article.
References
-
- Derégnaucourt S. Birdsong learning in the laboratory, with especial reference to the song of the Zebra Finch (Taeniopygia guttata) Interaction Studies. 2011;12:324–350.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
